Pazos Manuel, Casanova Mercedes, Palacios Pilar, Margolin William, Natale Paolo, Vicente Miguel
Centro Nacional de Biotecnología - Consejo Superior de Investigaciones Científicas (CNB-CSIC), Madrid, Spain.
Department of Microbiology and Molecular Genetics, University of Texas Medical School at Houston, Houston, Texas, United States of America.
PLoS One. 2014 Mar 17;9(3):e91984. doi: 10.1371/journal.pone.0091984. eCollection 2014.
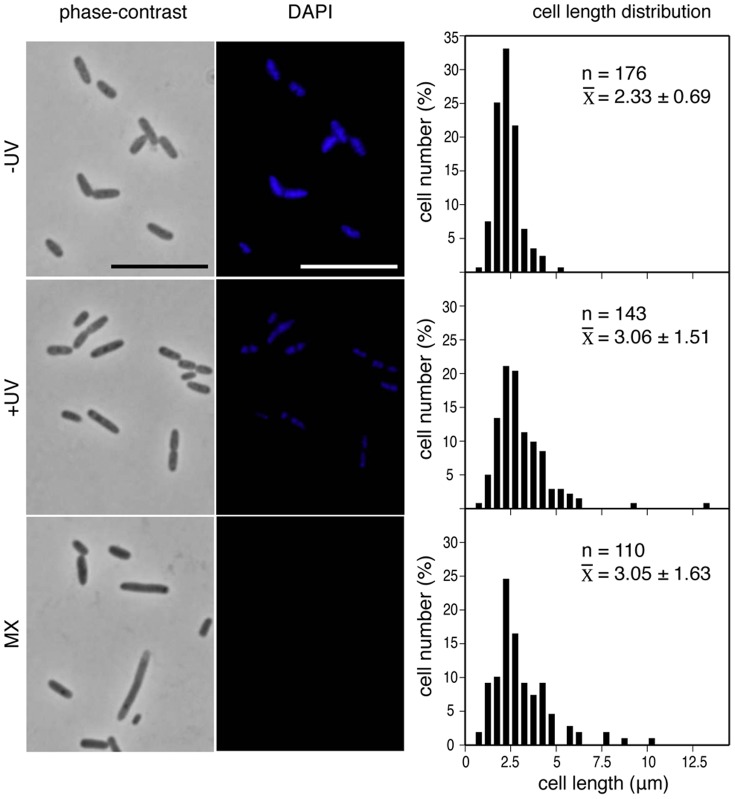
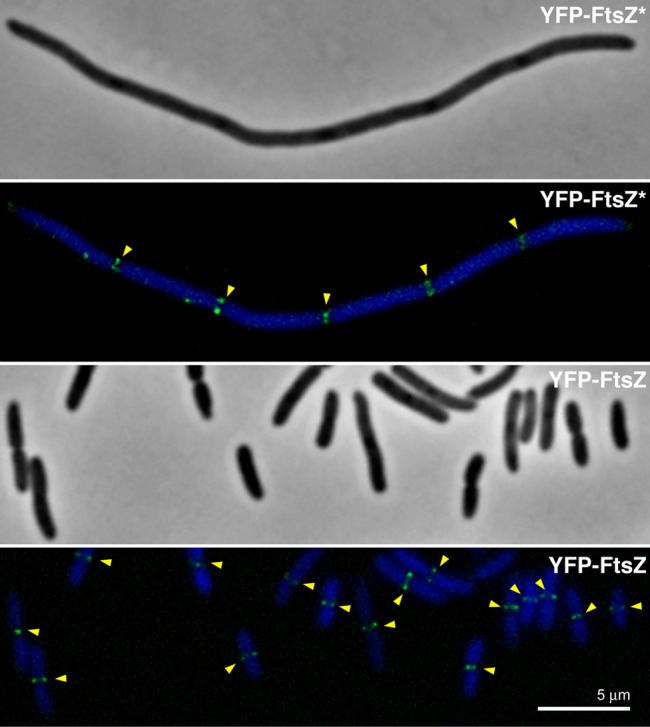
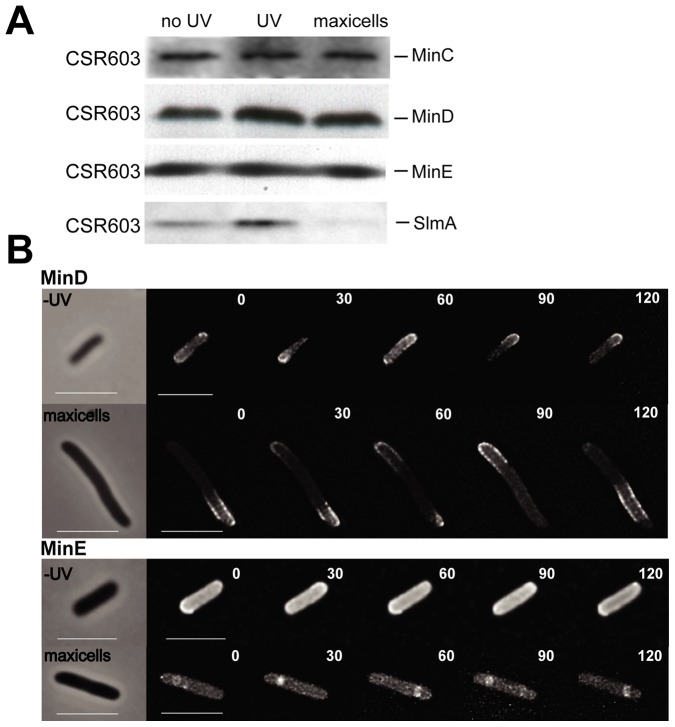
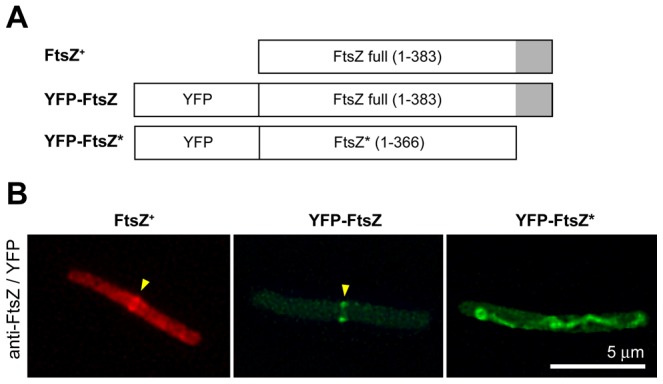
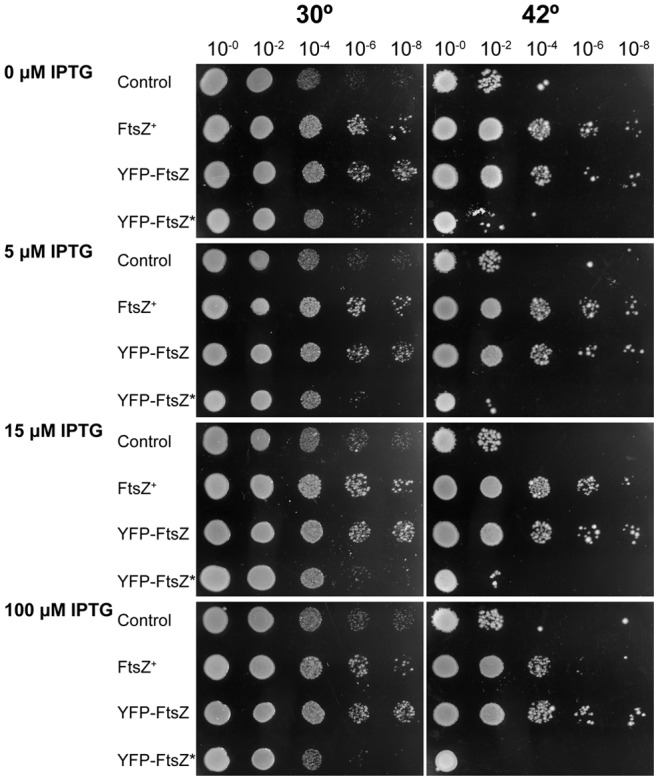
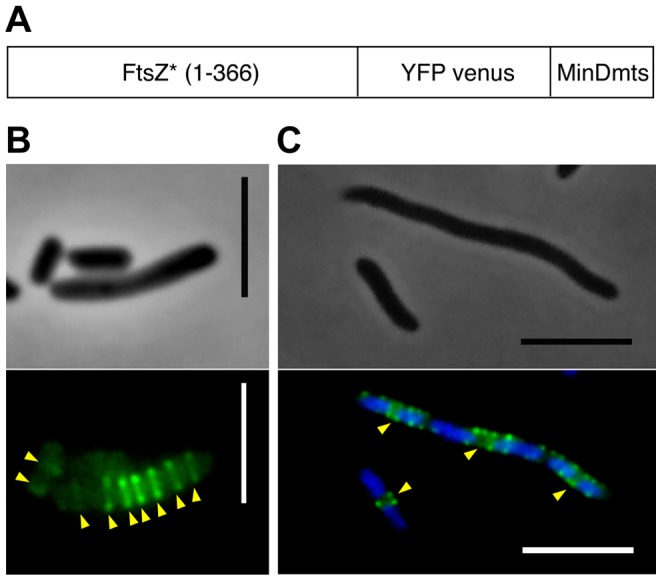
We describe the placement of the cytoplasmic FtsZ protein, an essential component of the division septum, in nucleoid-free Escherichia coli maxicells. The absence of the nucleoid is accompanied in maxicells by degradation of the SlmA protein. This protein, together with the nucleoid, prevents the placement of the septum in the regions occupied by the chromosome by a mechanism called nucleoid occlusion (NO). A second septum placement mechanism, the MinCDE system (Min) involving a pole-to-pole oscillation of three proteins, nonetheless remains active in maxicells. Both Min and NO act on the polymerization of FtsZ, preventing its assembly into an FtsZ-ring except at midcell. Our results show that even in the total absence of NO, Min oscillations can direct placement of FtsZ in maxicells. Deletion of the FtsZ carboxyl terminal domain (FtsZ*), a central hub that receives signals from a variety of proteins including MinC, FtsA and ZipA, produces a Min-insensitive form of FtsZ unable to interact with the membrane-anchoring FtsA and ZipA proteins. This protein produces a totally disorganized pattern of FtsZ localization inside the maxicell cytoplasm. In contrast, FtsZ*-VM, an artificially cytoplasmic membrane-anchored variant of FtsZ*, forms helical or repetitive ring structures distributed along the entire length of maxicells even in the absence of NO. These results show that membrane anchoring is needed to organize FtsZ into rings and underscore the role of the C-terminal hub of FtsZ for their correct placement.
我们描述了细胞质中FtsZ蛋白(分裂隔膜的一个重要组成部分)在无类核的大肠杆菌大细胞中的定位情况。在大细胞中,类核的缺失伴随着SlmA蛋白的降解。该蛋白与类核一起,通过一种称为类核阻隔(NO)的机制,防止隔膜在染色体占据的区域形成。第二种隔膜定位机制,即涉及三种蛋白极到极振荡的MinCDE系统(Min),在大细胞中仍然活跃。Min和NO都作用于FtsZ的聚合,阻止其组装成FtsZ环,除非在细胞中部。我们的结果表明,即使在完全没有NO的情况下,Min振荡也能指导FtsZ在大细胞中的定位。FtsZ羧基末端结构域(FtsZ*)的缺失,这是一个接收包括MinC、FtsA和ZipA等多种蛋白信号的中心枢纽,会产生一种对Min不敏感的FtsZ形式,无法与膜锚定蛋白FtsA和ZipA相互作用。这种蛋白在大细胞细胞质内产生完全紊乱的FtsZ定位模式。相比之下,FtsZ*-VM是一种人工构建的、锚定在细胞质膜上的FtsZ*变体,即使在没有NO的情况下,也能在大细胞的整个长度上形成螺旋状或重复的环状结构。这些结果表明,膜锚定是将FtsZ组织成环所必需的,并强调了FtsZ C末端枢纽对其正确定位的作用。