Bagai Ireena, Sarangi Ritimukta, Fleischhacker Angela S, Sharma Ajay, Hoffman Brian M, Zuiderweg Erik R P, Ragsdale Stephen W
†Department of Biological Chemistry, University of Michigan, Ann Arbor, Michigan 48019, United States.
‡Stanford Synchrotron Radiation Lightsource, SLAC National Accelerator Laboratory, Menlo Park, California 94025, United States.
Biochemistry. 2015 May 5;54(17):2693-708. doi: 10.1021/bi501489r. Epub 2015 Apr 22.
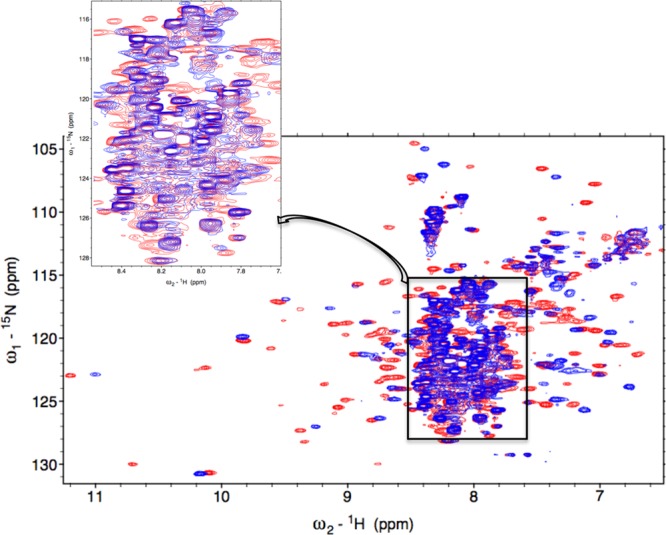
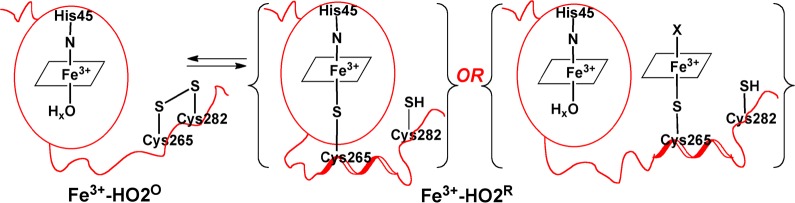
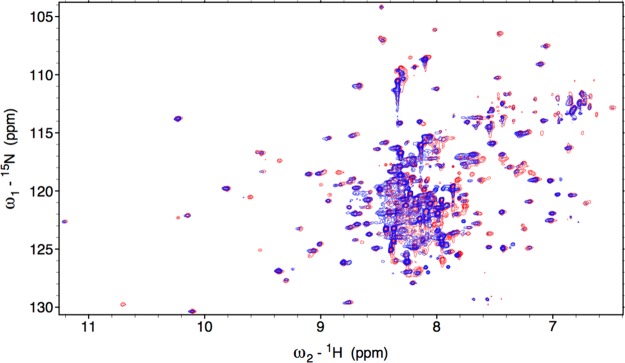
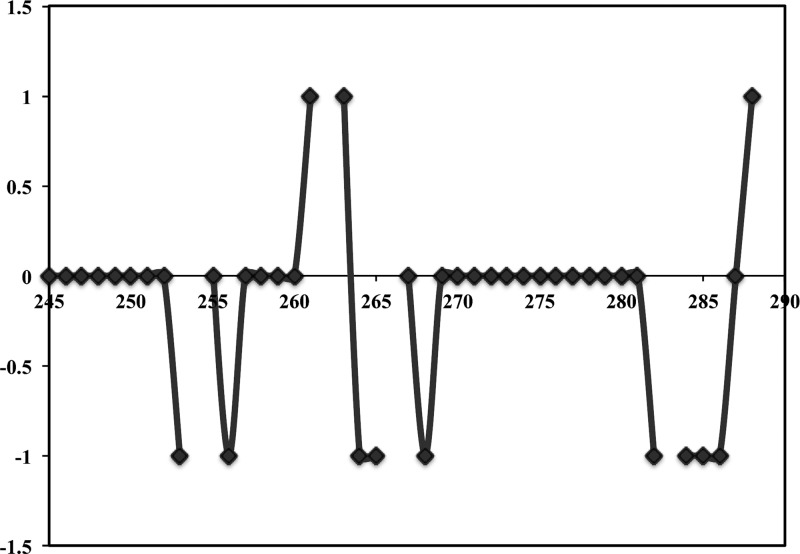
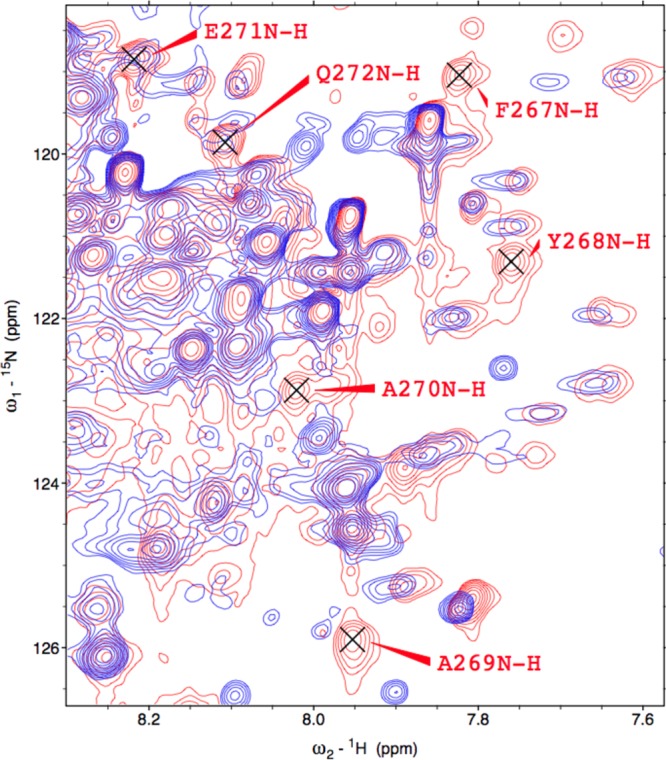
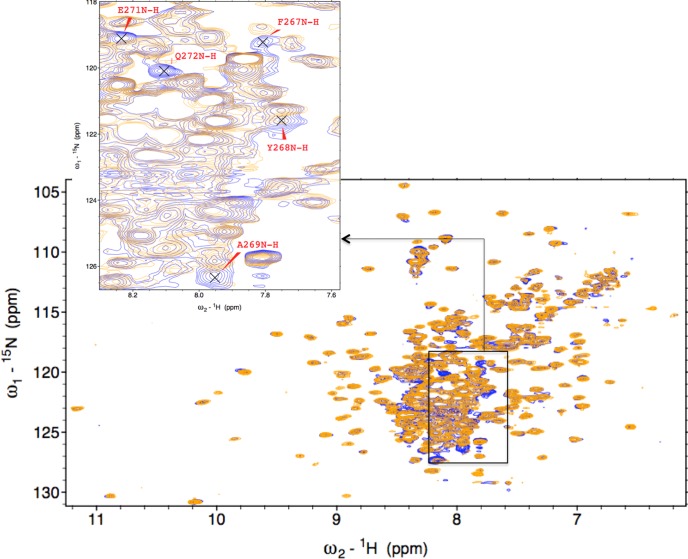
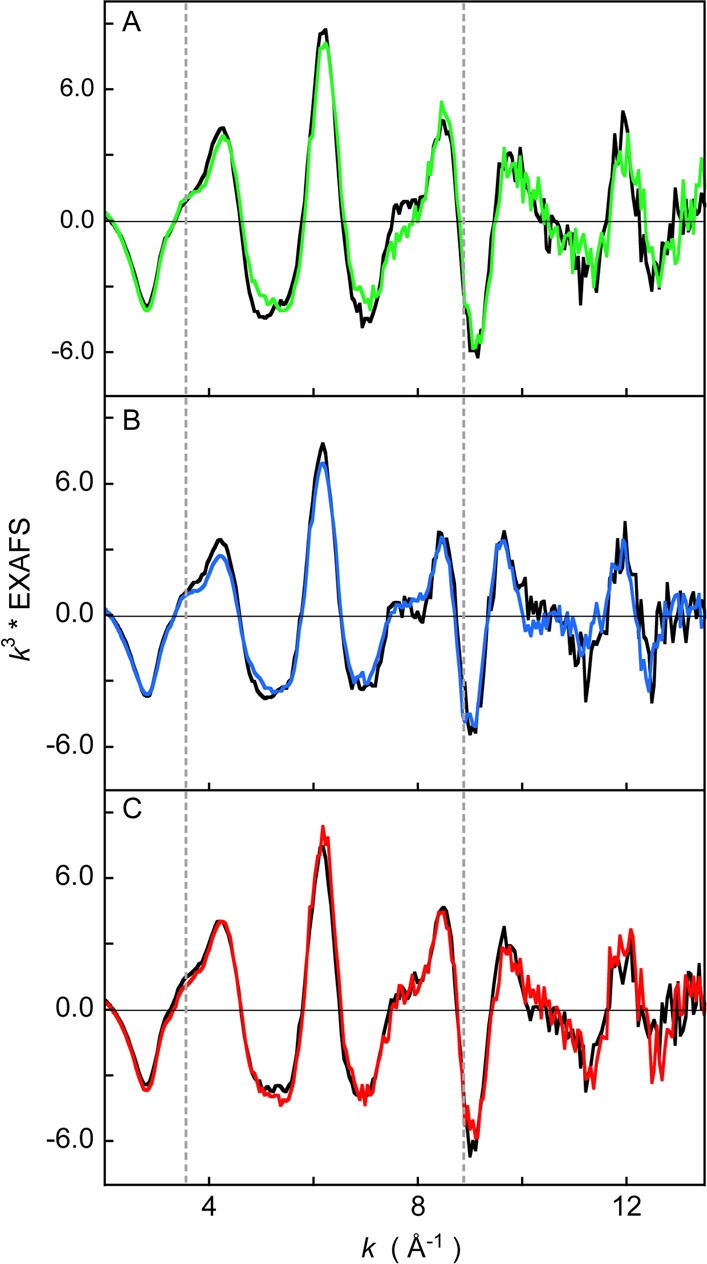
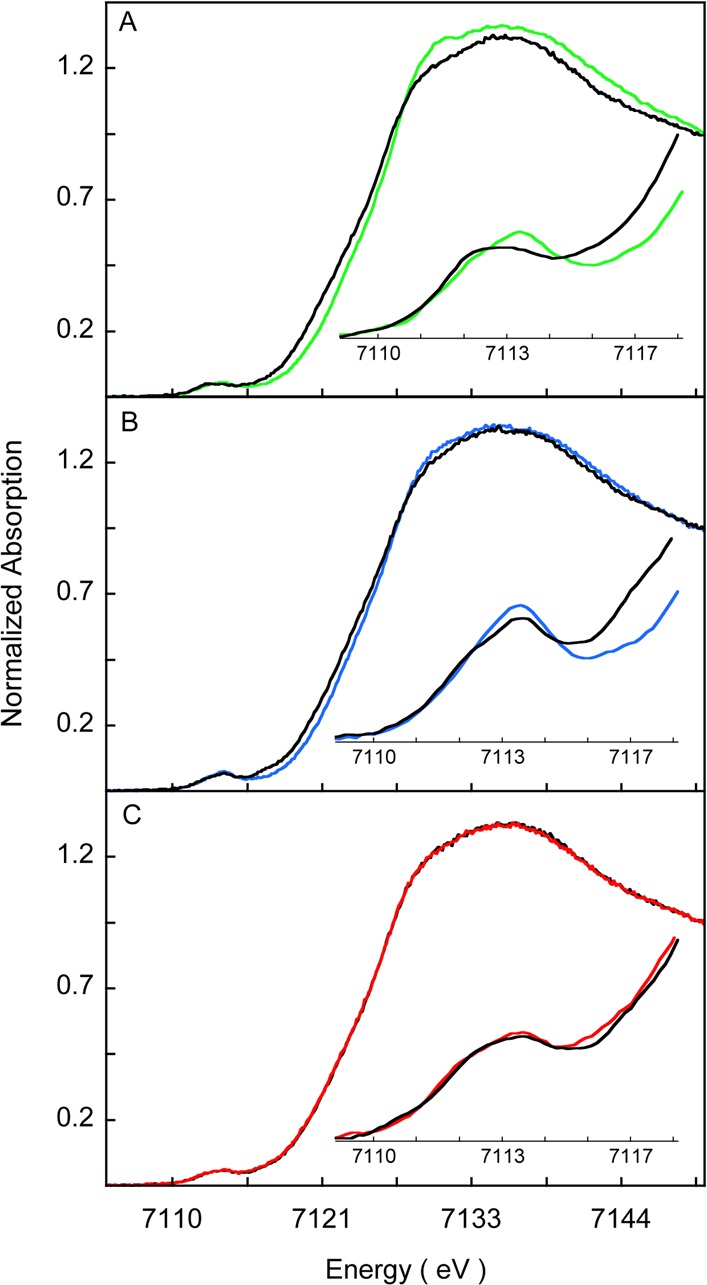
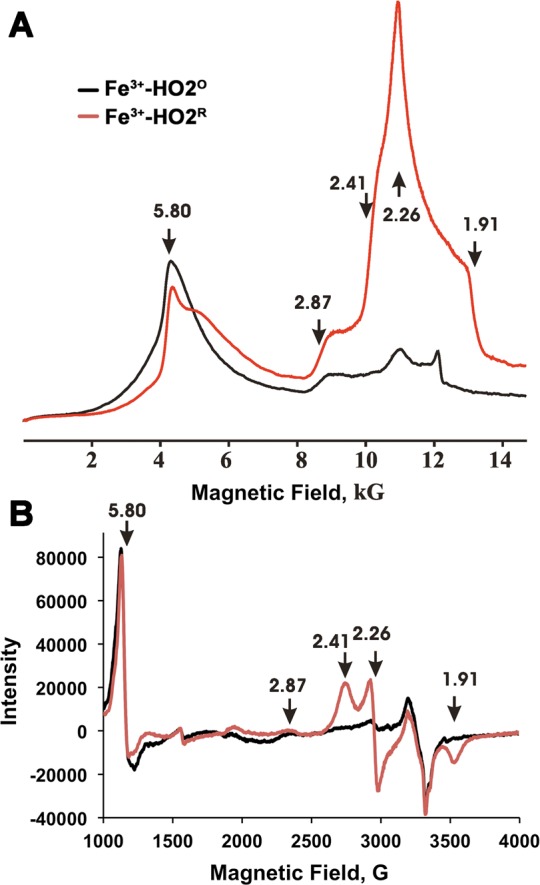
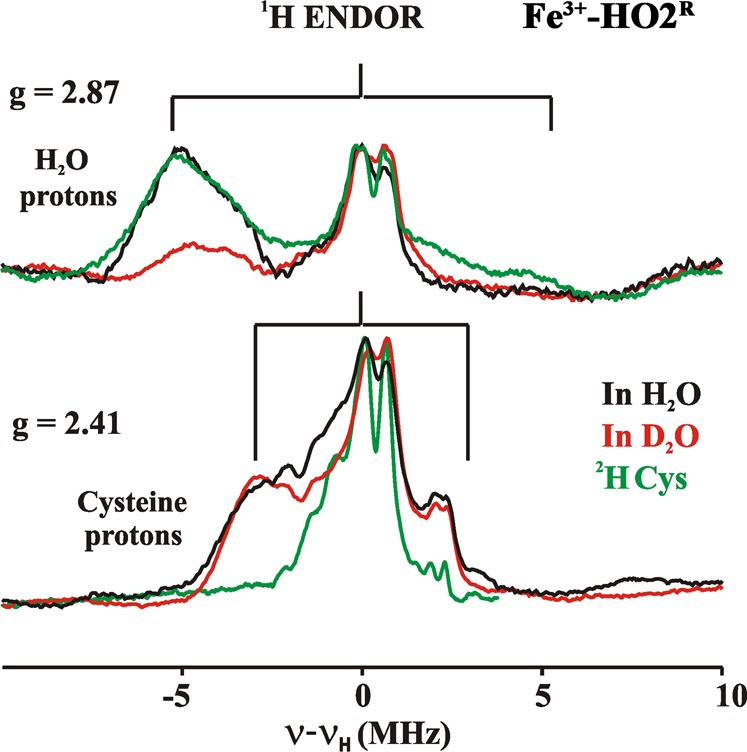
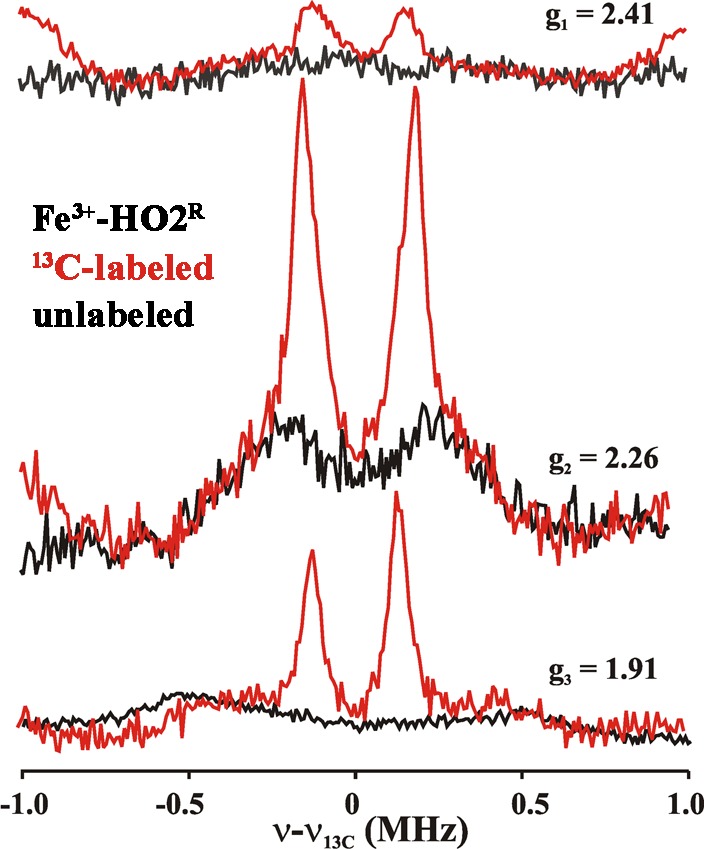
Heme oxygenase (HO) catalyzes a key step in heme homeostasis: the O2- and NADPH-cytochrome P450 reductase-dependent conversion of heme to biliverdin, Fe, and CO through a process in which the heme participates both as a prosthetic group and as a substrate. Mammals contain two isoforms of this enzyme, HO2 and HO1, which share the same α-helical fold forming the catalytic core and heme binding site, as well as a membrane spanning helix at their C-termini. However, unlike HO1, HO2 has an additional 30-residue N-terminus as well as two cysteine-proline sequences near the C-terminus that reside in heme regulatory motifs (HRMs). While the role of the additional N-terminal residues of HO2 is not yet understood, the HRMs have been proposed to reversibly form a thiol/disulfide redox switch that modulates the affinity of HO2 for ferric heme as a function of cellular redox poise. To further define the roles of the N- and C-terminal regions unique to HO2, we used multiple spectroscopic techniques to characterize these regions of the human HO2. Nuclear magnetic resonance spectroscopic experiments with HO2 demonstrate that, when the HRMs are in the oxidized state (HO2(O)), both the extra N-terminal and the C-terminal HRM-containing regions are disordered. However, protein NMR experiments illustrate that, under reducing conditions, the C-terminal region gains some structure as the Cys residues in the HRMs undergo reduction (HO2(R)) and, in experiments employing a diamagnetic protoporphyrin, suggest a redox-dependent interaction between the core and the HRM domains. Further, electron nuclear double resonance and X-ray absorption spectroscopic studies demonstrate that, upon reduction of the HRMs to the sulfhydryl form, a cysteine residue from the HRM region ligates to a ferric heme. Taken together with EPR measurements, which show the appearance of a new low-spin heme signal in reduced HO2, it appears that a cysteine residue(s) in the HRMs directly interacts with a second bound heme.
血红素加氧酶(HO)催化血红素稳态中的关键步骤:通过一个血红素既作为辅基又作为底物参与的过程,将血红素在氧气和NADPH - 细胞色素P450还原酶的依赖下转化为胆绿素、铁和一氧化碳。哺乳动物含有该酶的两种同工型,即HO2和HO1,它们具有相同的形成催化核心和血红素结合位点的α - 螺旋折叠,以及在其C末端的一个跨膜螺旋。然而,与HO1不同,HO2有一个额外的30个残基的N末端,以及在C末端附近的两个半胱氨酸 - 脯氨酸序列,它们位于血红素调节基序(HRM)中。虽然HO2额外的N末端残基的作用尚不清楚,但有人提出HRM可逆地形成一个硫醇/二硫键氧化还原开关,该开关根据细胞氧化还原状态调节HO2对三价铁血红素的亲和力。为了进一步确定HO2特有的N末端和C末端区域的作用,我们使用了多种光谱技术来表征人HO2的这些区域。对HO2进行的核磁共振光谱实验表明,当HRM处于氧化状态(HO2(O))时,额外的N末端和含C末端HRM的区域都是无序的。然而,蛋白质NMR实验表明,在还原条件下,随着HRM中的半胱氨酸残基发生还原(HO2(R)),C末端区域获得了一些结构,并且在使用抗磁性原卟啉的实验中,表明核心和HRM结构域之间存在氧化还原依赖性相互作用。此外,电子核双共振和X射线吸收光谱研究表明,当HRM还原为巯基形式时,HRM区域的一个半胱氨酸残基与三价铁血红素结合。结合电子顺磁共振测量结果,该结果显示在还原的HO2中出现了一个新的低自旋血红素信号,似乎HRM中的一个或多个半胱氨酸残基直接与第二个结合的血红素相互作用。