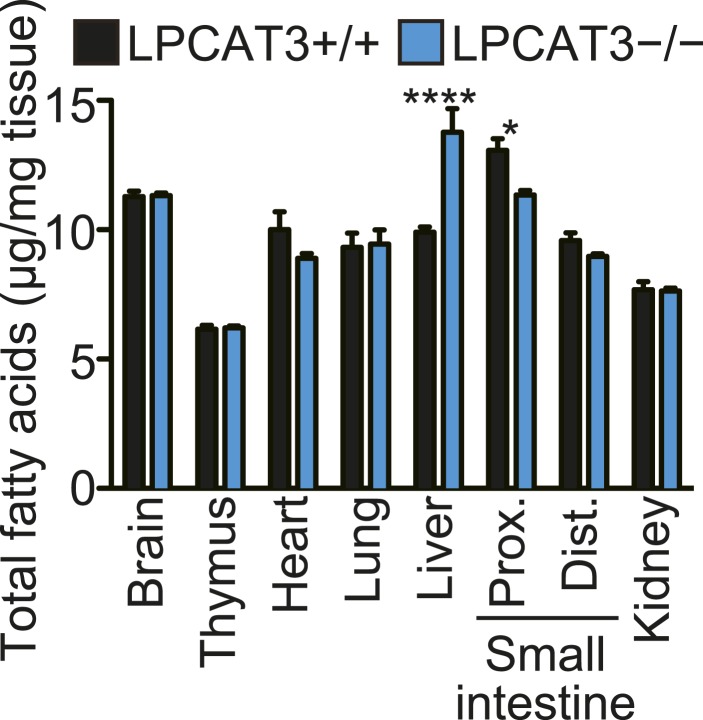
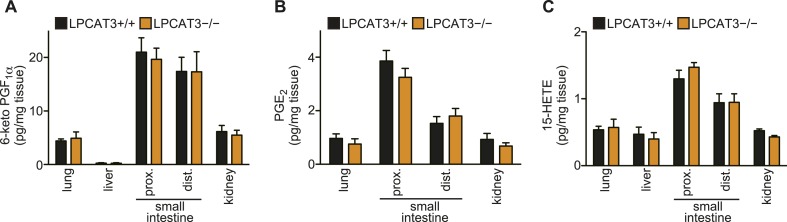
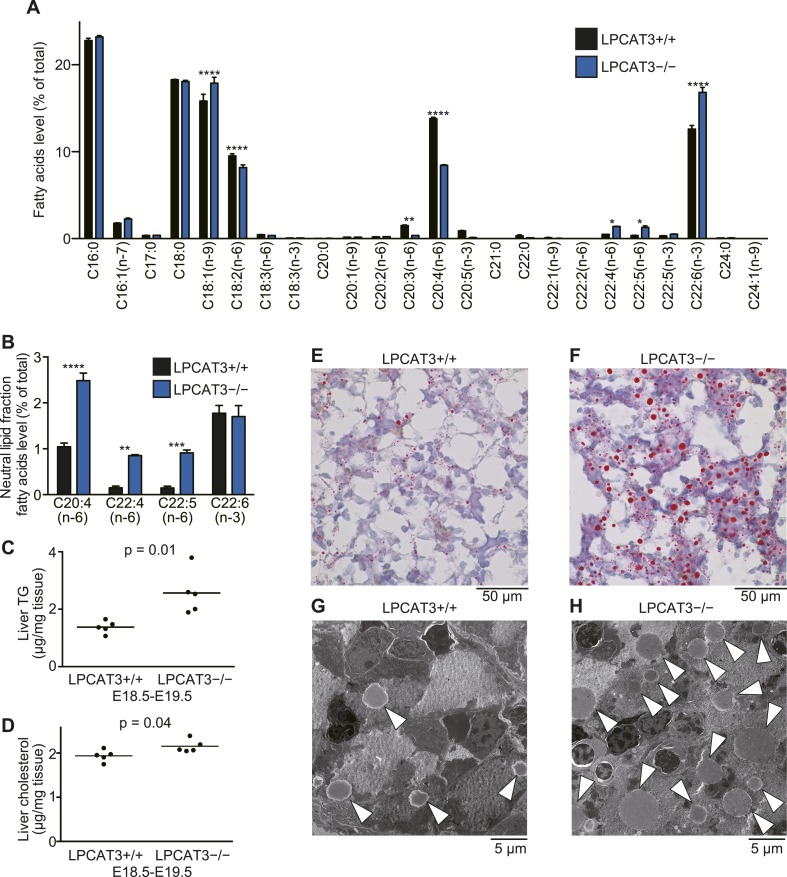
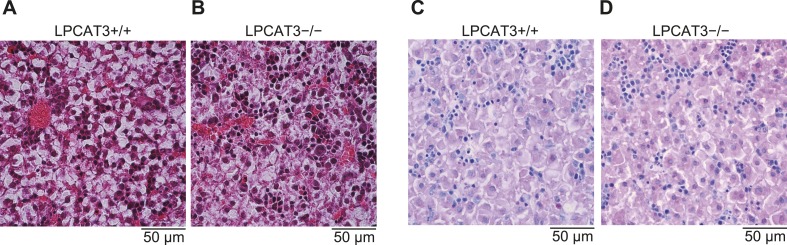
Hashidate-Yoshida Tomomi, Harayama Takeshi, Hishikawa Daisuke, Morimoto Ryo, Hamano Fumie, Tokuoka Suzumi M, Eto Miki, Tamura-Nakano Miwa, Yanobu-Takanashi Rieko, Mukumoto Yoshiko, Kiyonari Hiroshi, Okamura Tadashi, Kita Yoshihiro, Shindou Hideo, Shimizu Takao
Department of Lipid Signaling, National Center for Global Health and Medicine, Tokyo, Japan.
Department of Lipidomics, Graduate School of Medicine, The University of Tokyo, Tokyo, Japan.
Elife. 2015 Apr 21;4:e06328. doi: 10.7554/eLife.06328.
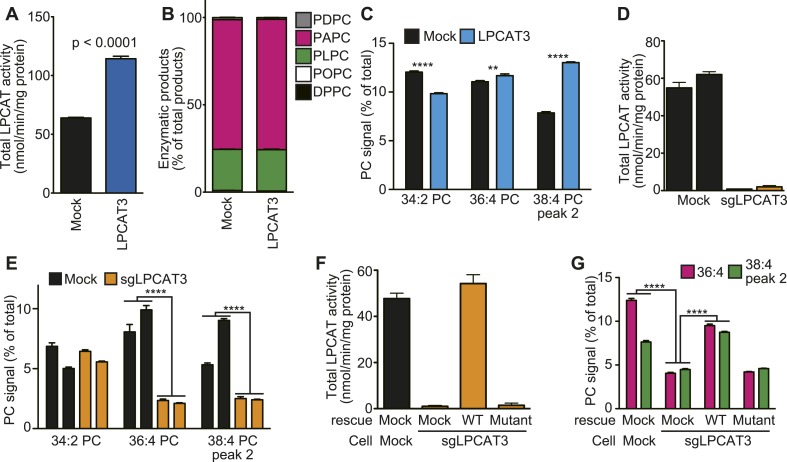
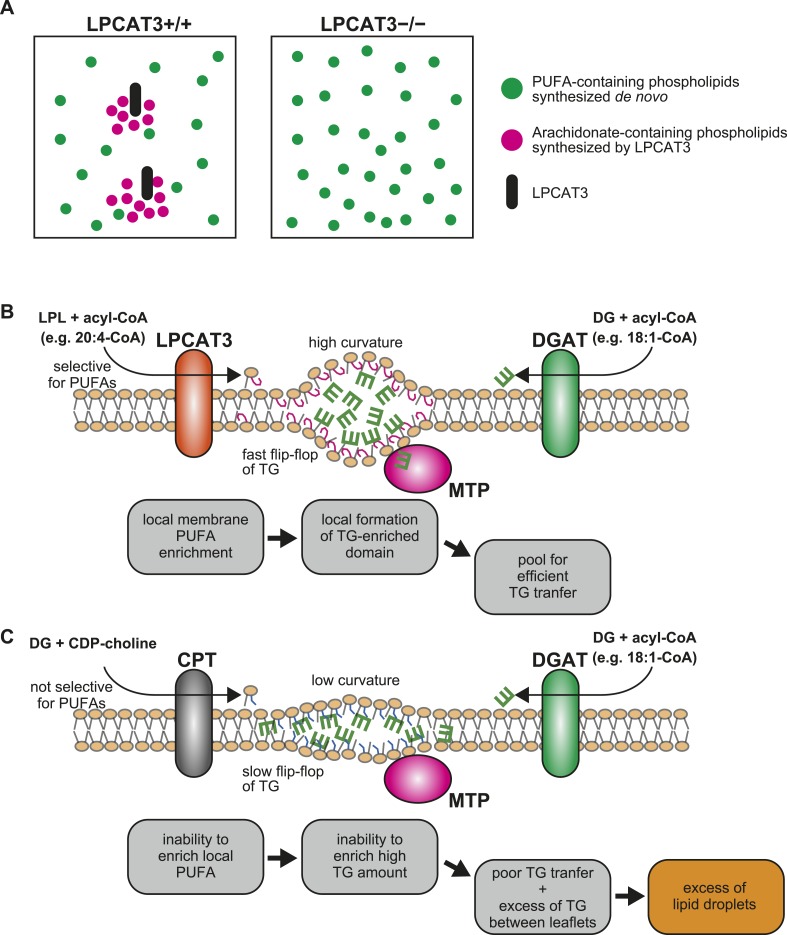
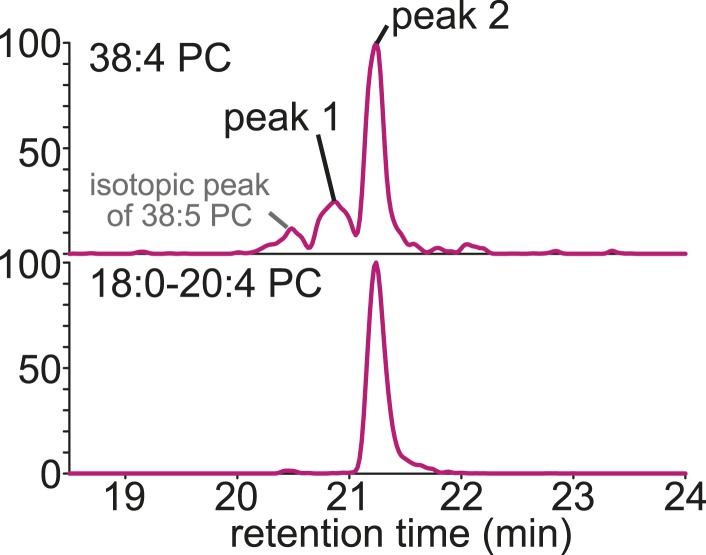
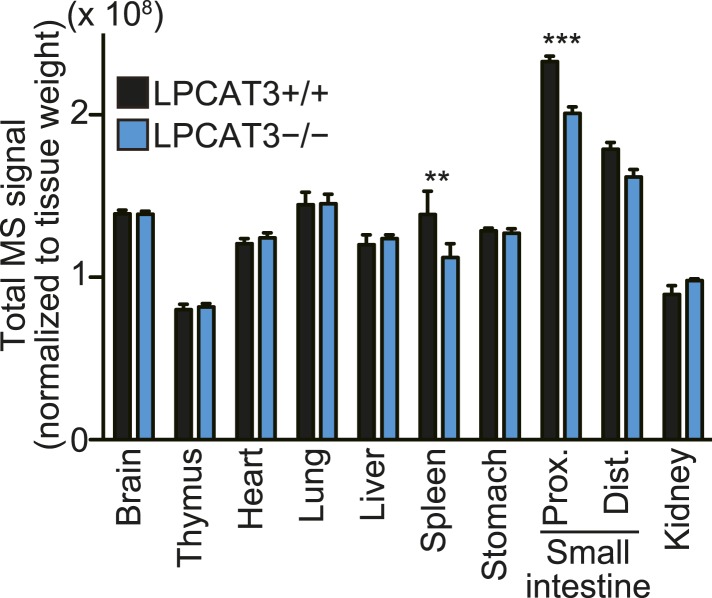
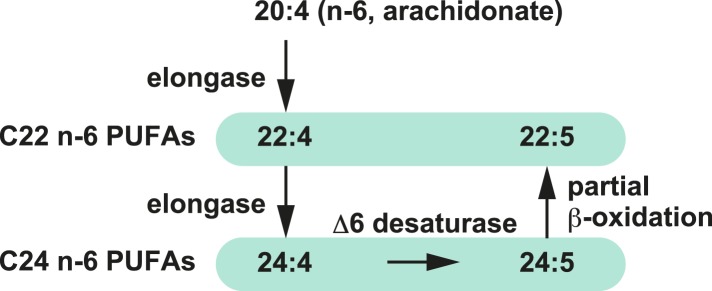
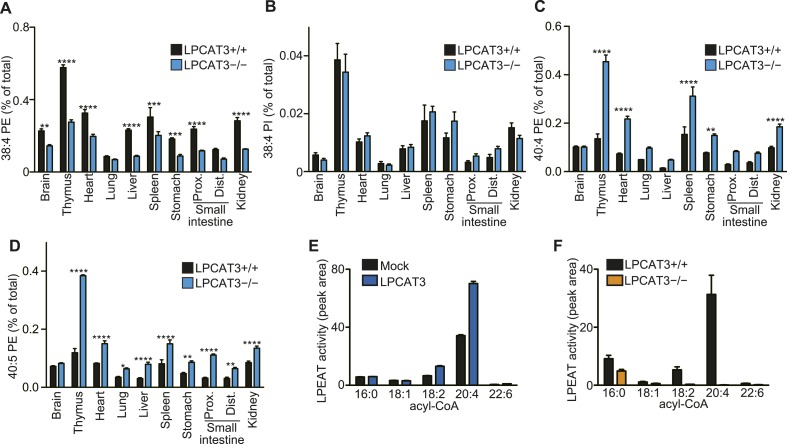
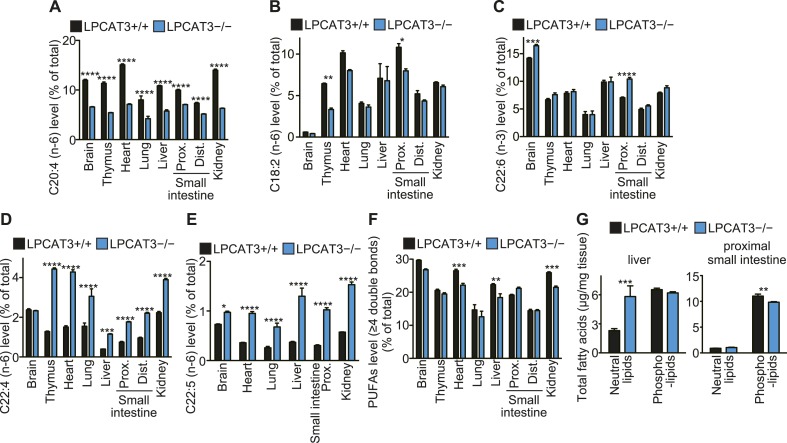
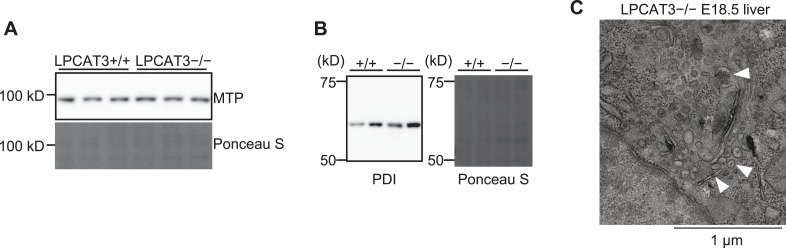
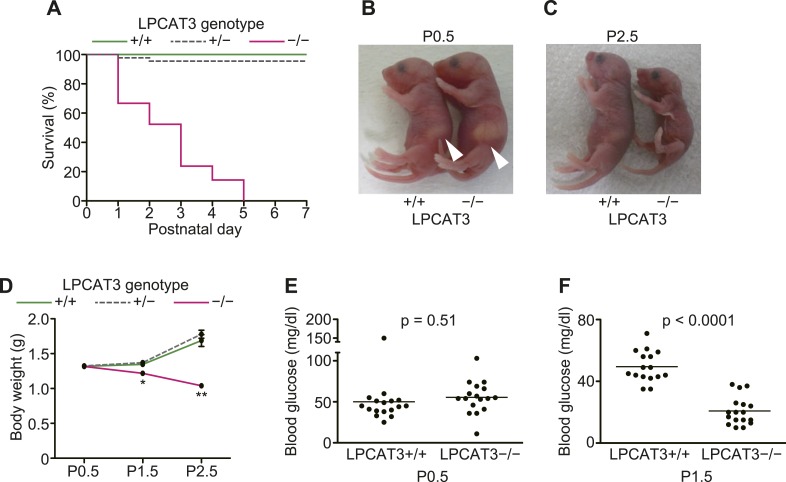
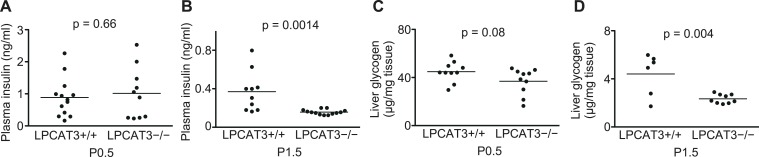
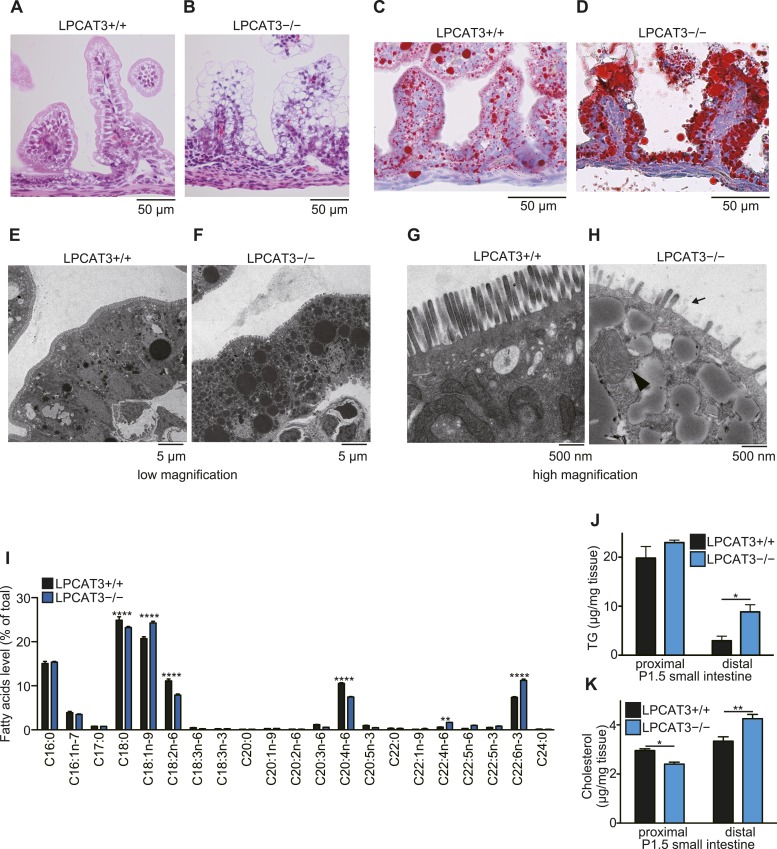
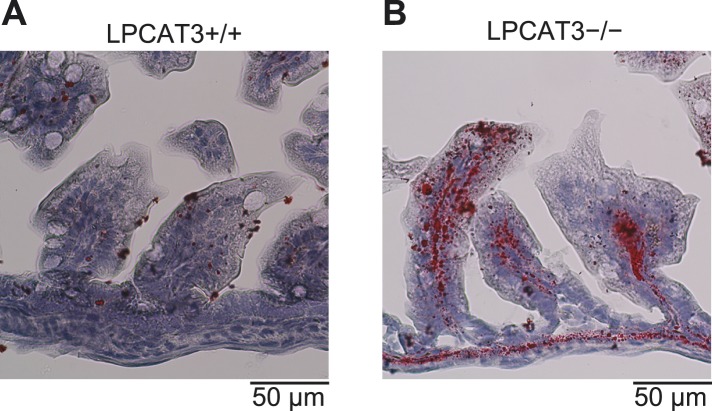
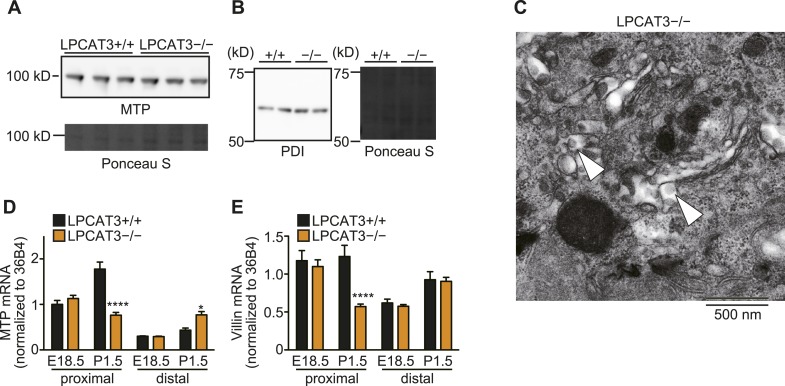
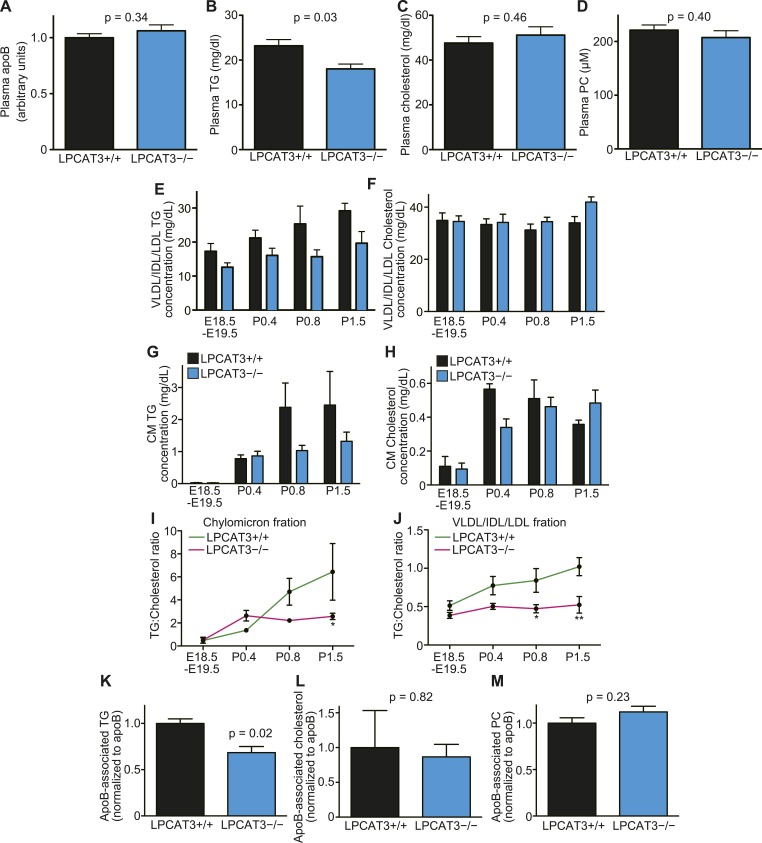
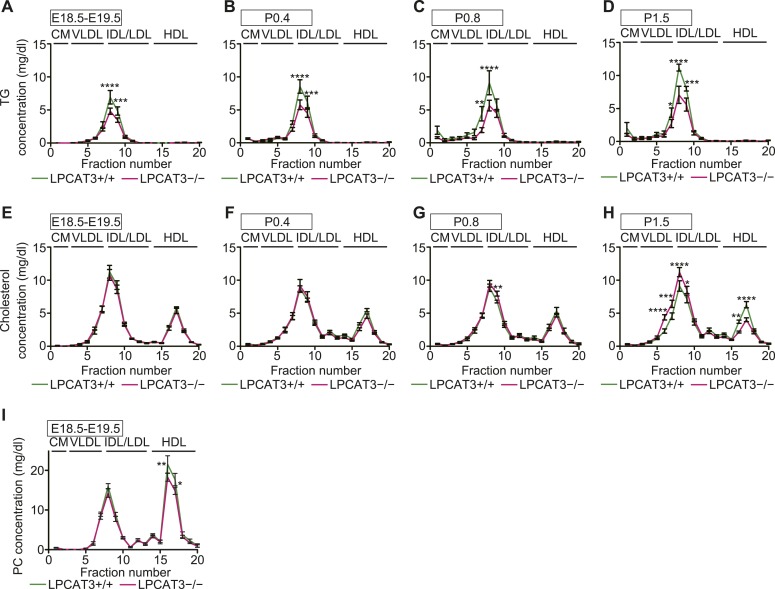
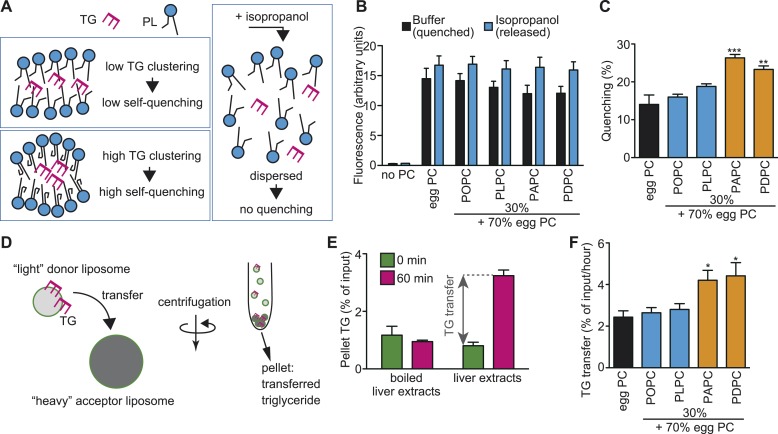
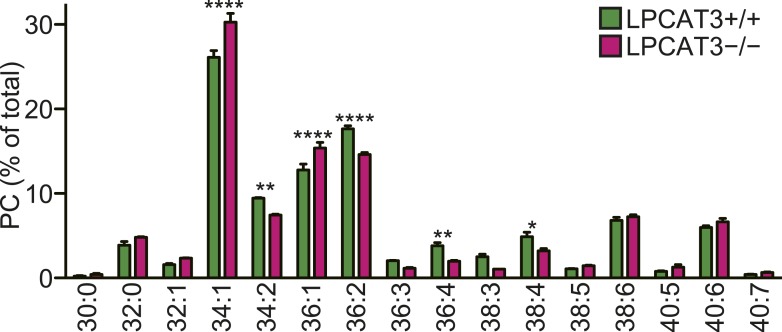
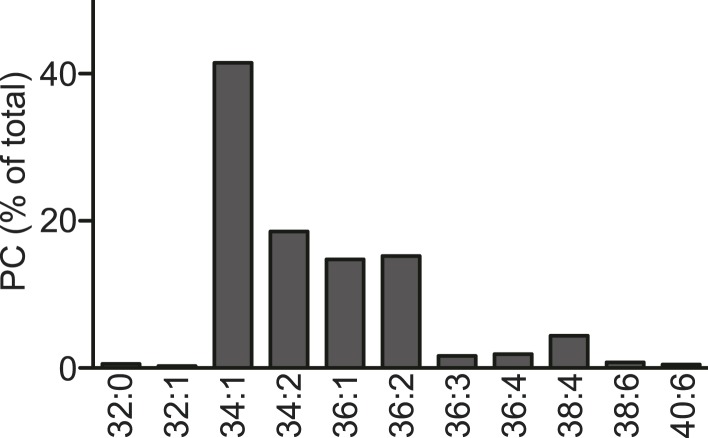
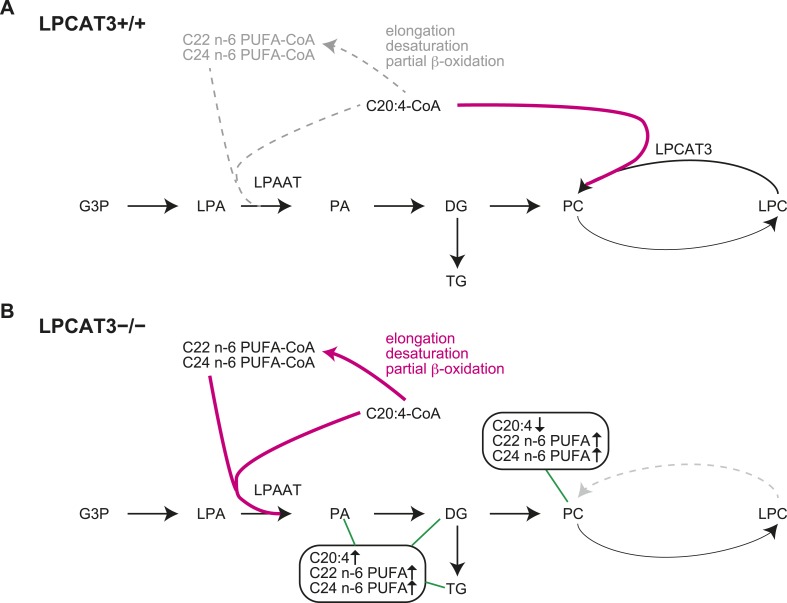
Polyunsaturated fatty acids (PUFAs) in phospholipids affect the physical properties of membranes, but it is unclear which biological processes are influenced by their regulation. For example, the functions of membrane arachidonate that are independent of a precursor role for eicosanoid synthesis remain largely unknown. Here, we show that the lack of lysophosphatidylcholine acyltransferase 3 (LPCAT3) leads to drastic reductions in membrane arachidonate levels, and that LPCAT3-deficient mice are neonatally lethal due to an extensive triacylglycerol (TG) accumulation and dysfunction in enterocytes. We found that high levels of PUFAs in membranes enable TGs to locally cluster in high density, and that this clustering promotes efficient TG transfer. We propose a model of local arachidonate enrichment by LPCAT3 to generate a distinct pool of TG in membranes, which is required for normal directionality of TG transfer and lipoprotein assembly in the liver and enterocytes.
磷脂中的多不饱和脂肪酸(PUFAs)会影响细胞膜的物理性质,但尚不清楚它们的调控会影响哪些生物学过程。例如,膜花生四烯酸独立于类花生酸合成前体作用的功能在很大程度上仍不为人知。在此,我们表明溶血磷脂酰胆碱酰基转移酶3(LPCAT3)的缺失会导致膜花生四烯酸水平急剧降低,并且LPCAT3缺陷型小鼠在新生期致死,原因是肠细胞中大量三酰甘油(TG)积累和功能障碍。我们发现细胞膜中高水平的PUFAs能使TG在局部高密度聚集,并且这种聚集促进了TG的高效转移。我们提出了一个由LPCAT3进行局部花生四烯酸富集以在细胞膜中生成不同TG池的模型,这是肝脏和肠细胞中TG转移和脂蛋白组装正常方向性所必需的。