Alswied Abdullah, Parekh Anant B
From the Department of Physiology, Anatomy, and Genetics, University of Oxford, Oxford OX1 3PT, United Kingdom.
From the Department of Physiology, Anatomy, and Genetics, University of Oxford, Oxford OX1 3PT, United Kingdom
J Biol Chem. 2015 Dec 4;290(49):29555-66. doi: 10.1074/jbc.M115.678292. Epub 2015 Oct 14.
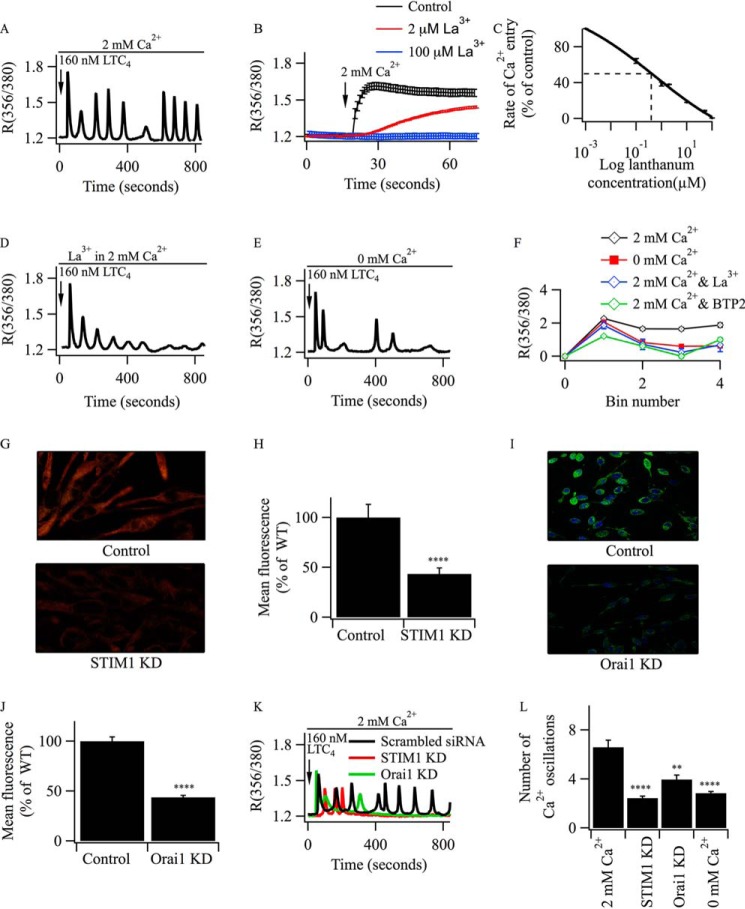
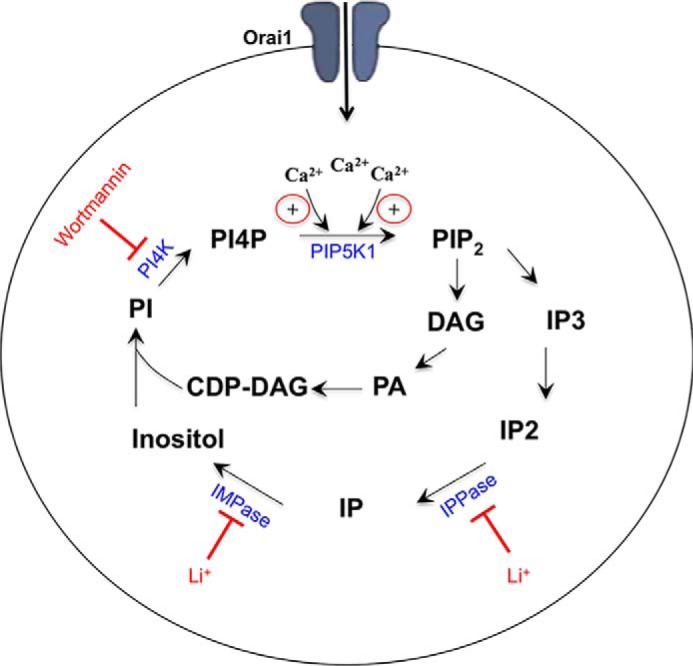
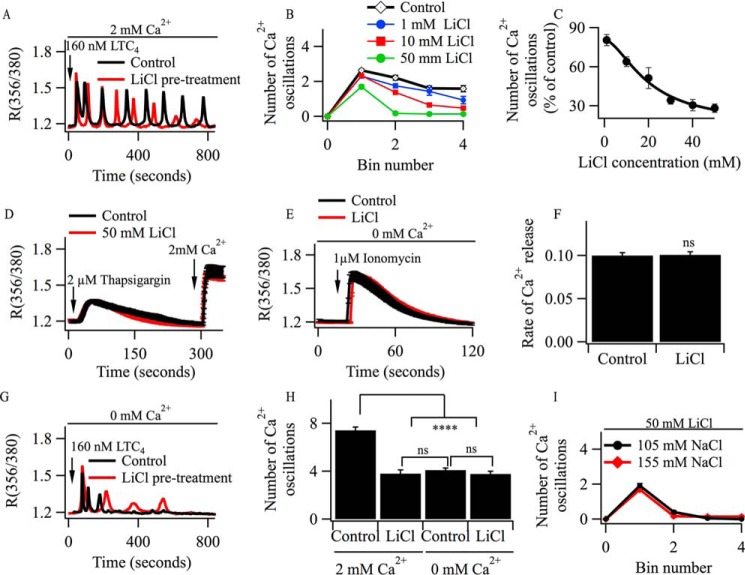
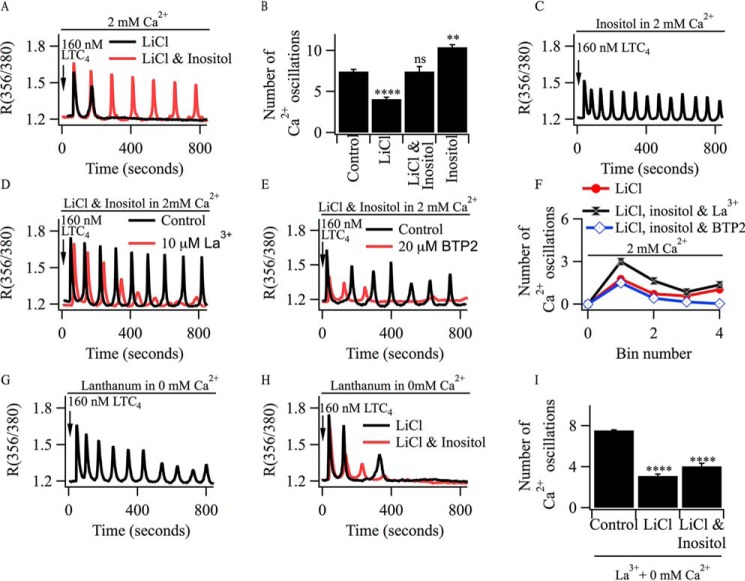
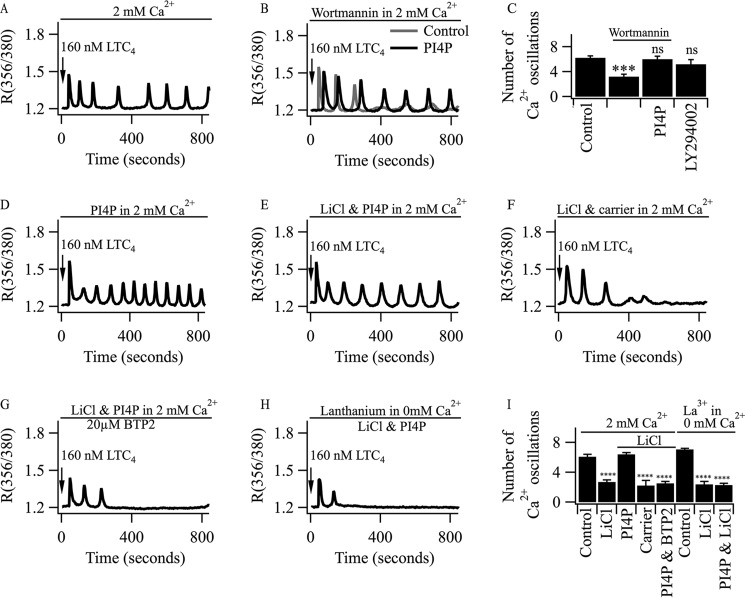
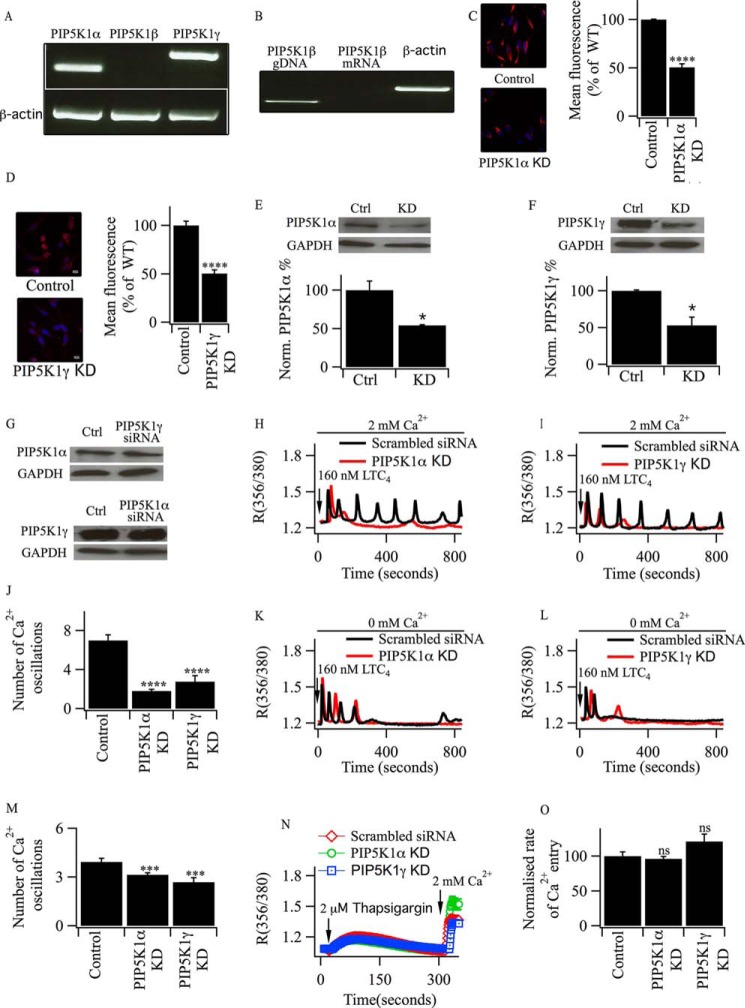
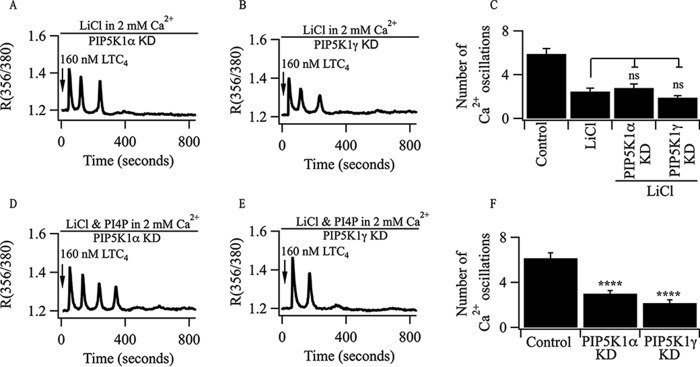
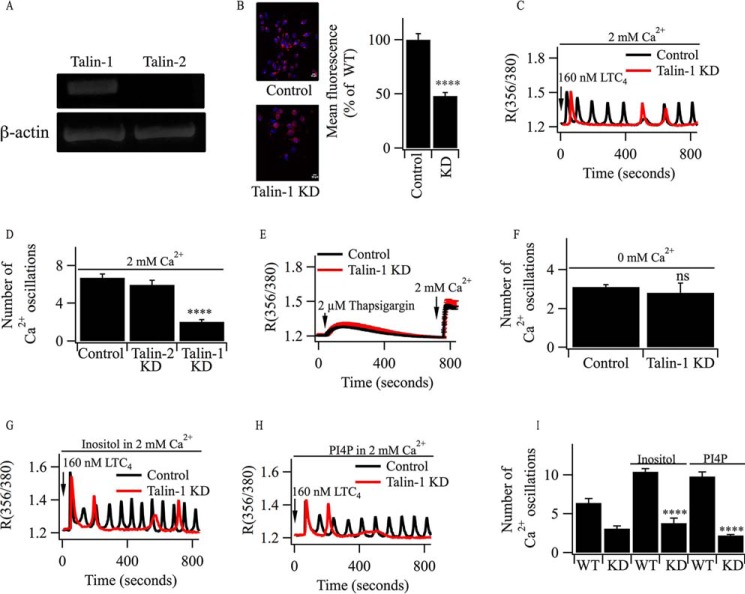
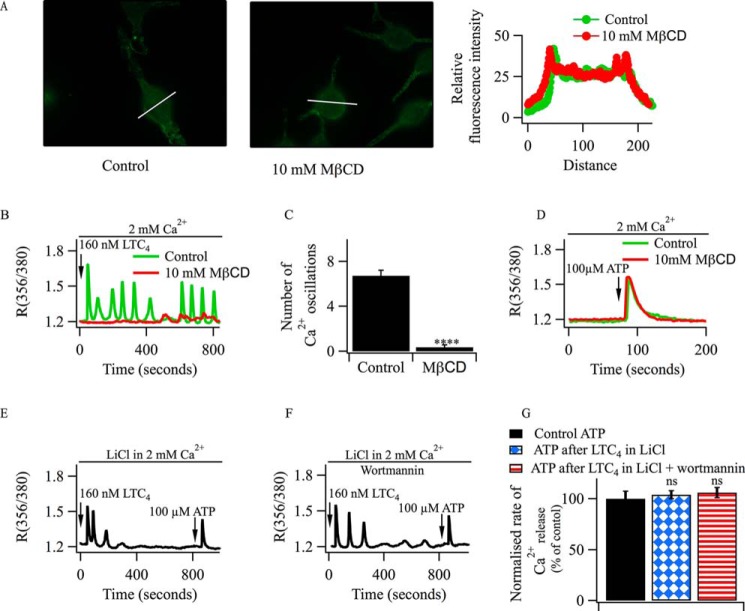
Oscillations in cytoplasmic Ca(2+) concentration are a universal mode of signaling following physiological levels of stimulation with agonists that engage the phospholipase C pathway. Sustained cytoplasmic Ca(2+) oscillations require replenishment of the membrane phospholipid phosphatidylinositol 4,5-bisphosphate (PIP2), the source of the Ca(2+)-releasing second messenger inositol trisphosphate. Here we show that cytoplasmic Ca(2+) oscillations induced by cysteinyl leukotriene type I receptor activation run down when cells are pretreated with Li(+), an inhibitor of inositol monophosphatases that prevents PIP2 resynthesis. In Li(+)-treated cells, cytoplasmic Ca(2+) signals evoked by an agonist were rescued by addition of exogenous inositol or phosphatidylinositol 4-phosphate (PI4P). Knockdown of the phosphatidylinositol 4-phosphate 5 (PIP5) kinases α and γ resulted in rapid loss of the intracellular Ca(2+) oscillations and also prevented rescue by PI4P. Knockdown of talin1, a protein that helps regulate PIP5 kinases, accelerated rundown of cytoplasmic Ca(2+) oscillations, and these could not be rescued by inositol or PI4P. In Li(+)-treated cells, recovery of the cytoplasmic Ca(2+) oscillations in the presence of inositol or PI4P was suppressed when Ca(2+) influx through store-operated Ca(2+) channels was inhibited. After rundown of the Ca(2+) signals following leukotriene receptor activation, stimulation of P2Y receptors evoked prominent inositol trisphosphate-dependent Ca(2+) release. Therefore, leukotriene and P2Y receptors utilize distinct membrane PIP2 pools. Our findings show that store-operated Ca(2+) entry is needed to sustain cytoplasmic Ca(2+) signaling following leukotriene receptor activation both by refilling the Ca(2+) stores and by helping to replenish the PIP2 pool accessible to leukotriene receptors, ostensibly through control of PIP5 kinase activity.
细胞质中钙离子(Ca(2+))浓度的振荡是在通过激活磷脂酶C途径的激动剂进行生理水平刺激后普遍存在的信号传导模式。持续的细胞质Ca(2+)振荡需要补充膜磷脂磷脂酰肌醇4,5 - 二磷酸(PIP2),它是释放Ca(2+)的第二信使肌醇三磷酸的来源。在这里我们表明,当细胞用Li(+)预处理时,由I型半胱氨酰白三烯受体激活诱导的细胞质Ca(2+)振荡会减弱,Li(+)是肌醇单磷酸酶的抑制剂,可阻止PIP2的重新合成。在用Li(+)处理的细胞中,通过添加外源性肌醇或磷脂酰肌醇4 - 磷酸(PI4P)可挽救由激动剂引起的细胞质Ca(2+)信号。磷脂酰肌醇4 - 磷酸5(PIP5)激酶α和γ的敲低导致细胞内Ca(2+)振荡迅速丧失,并且也阻止了PI4P的挽救作用。talin1是一种有助于调节PIP5激酶的蛋白质,其敲低加速了细胞质Ca(2+)振荡的减弱,并且这些振荡不能被肌醇或PI4P挽救。在用Li(+)处理的细胞中,当通过储存 - 操纵性Ca(2+)通道的Ca(2+)内流被抑制时,在存在肌醇或PI4P的情况下细胞质Ca(2+)振荡的恢复受到抑制。在白三烯受体激活后Ca(2+)信号减弱后,刺激P2Y受体可引起显著的肌醇三磷酸依赖性Ca(2+)释放。因此,白三烯和P2Y受体利用不同的膜PIP2池。我们的研究结果表明,储存 - 操纵性Ca(2+)内流对于在白三烯受体激活后维持细胞质Ca(2+)信号传导是必需的,这既通过重新填充Ca(2+)储存,也通过帮助补充白三烯受体可利用的PIP2池,表面上是通过控制PIP5激酶活性来实现的。