Luque Niceto R, Garrido Jesús A, Naveros Francisco, Carrillo Richard R, D'Angelo Egidio, Ros Eduardo
Department of Computer Architecture and Technology, Research Centre for Information and Communications Technologies of the University of Granada (CITIC-UGR) Granada, Spain.
Brain Connectivity Center, Istituto di Ricovero e Cura a Carattere Scientifico, Istituto Neurologico Nazionale Casimiro MondinoPavia, Italy; Department of Brain and Behavioural Sciences, University of PaviaPavia, Italy.
Front Comput Neurosci. 2016 Mar 2;10:17. doi: 10.3389/fncom.2016.00017. eCollection 2016.
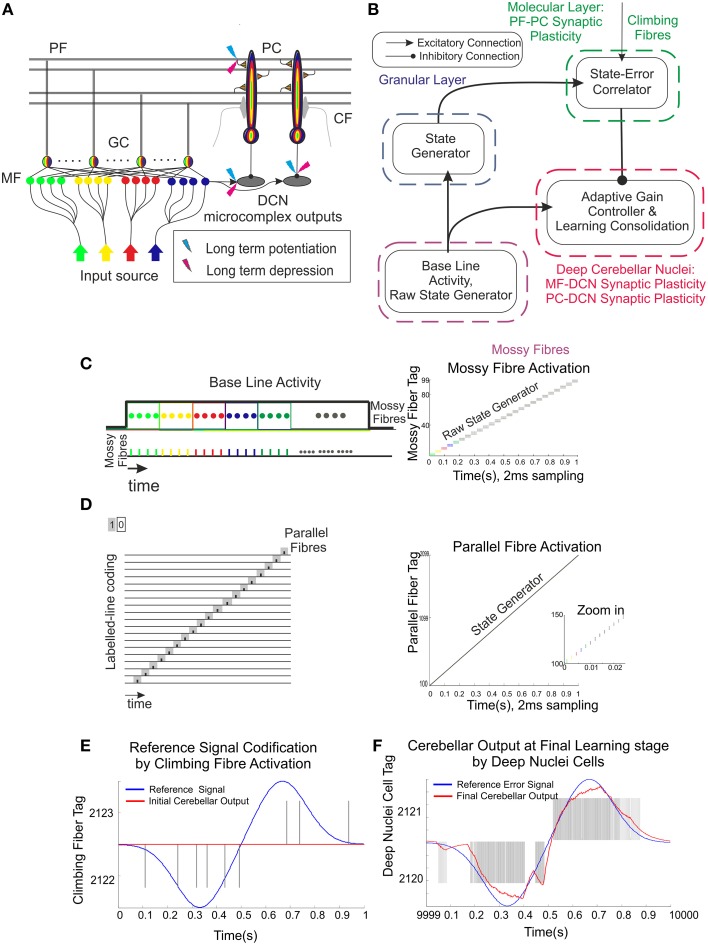
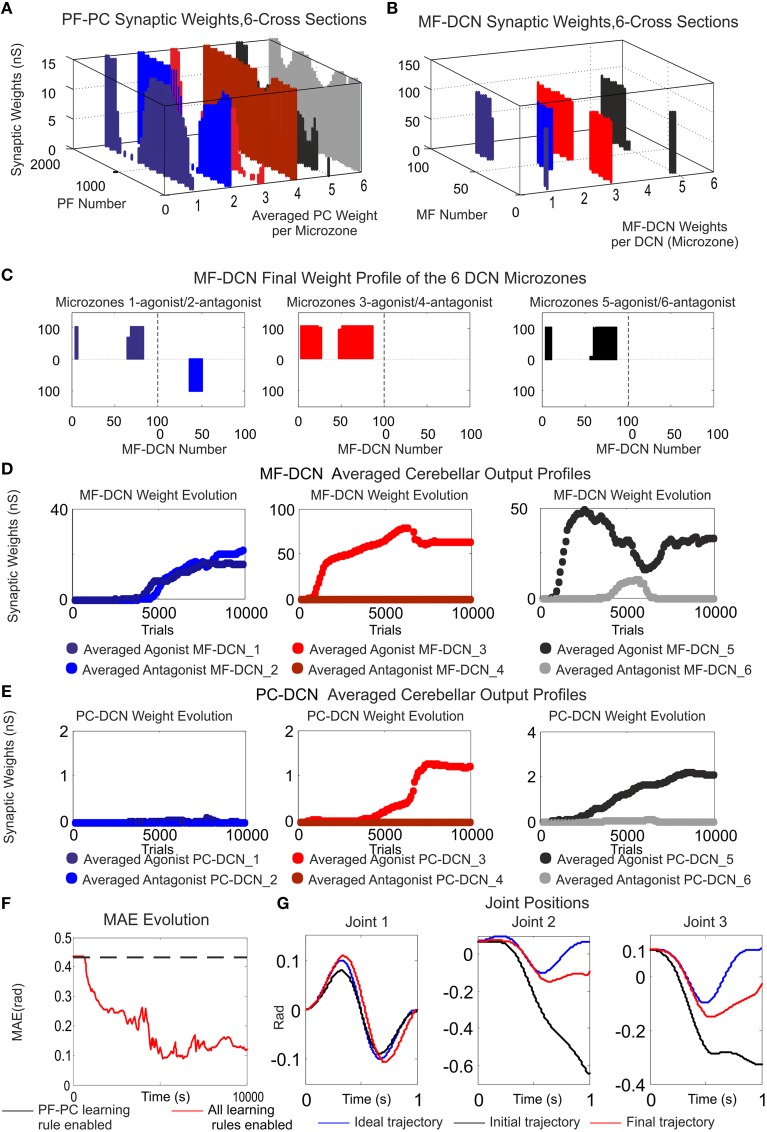
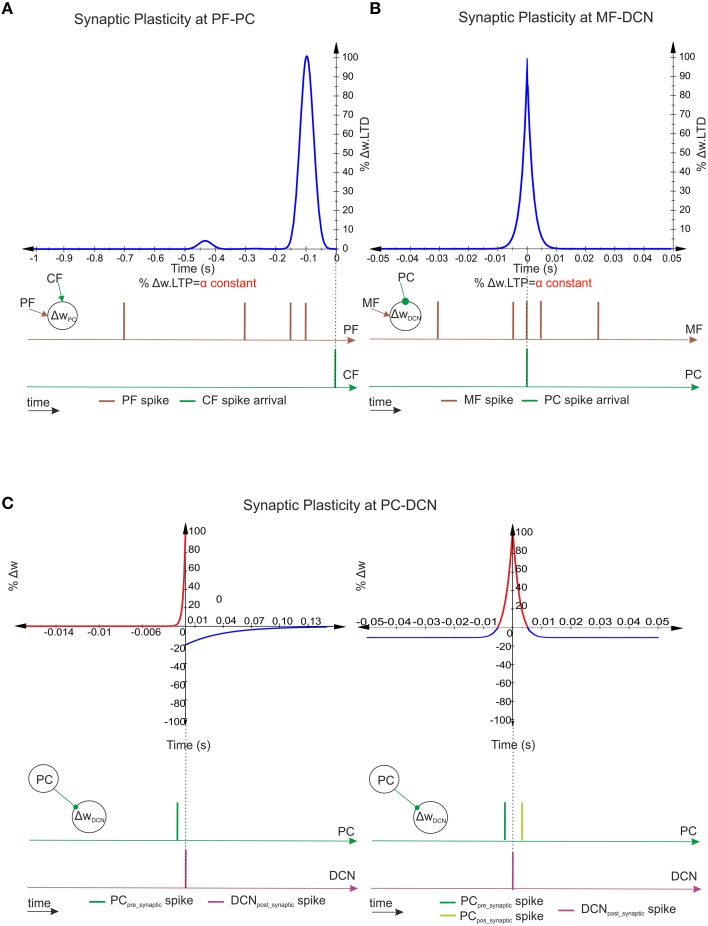
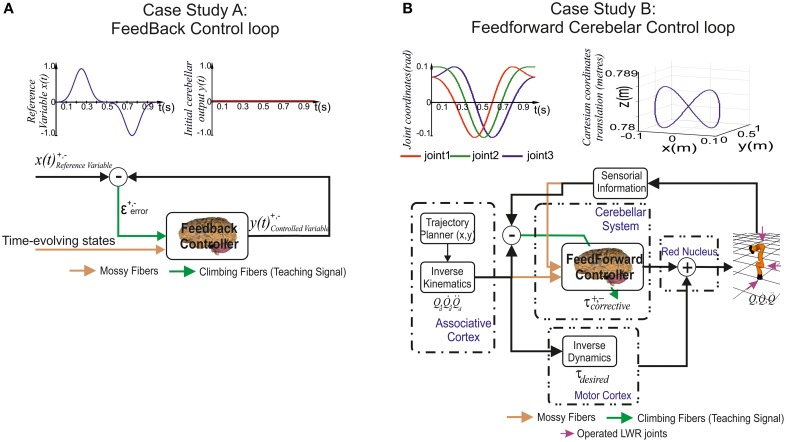
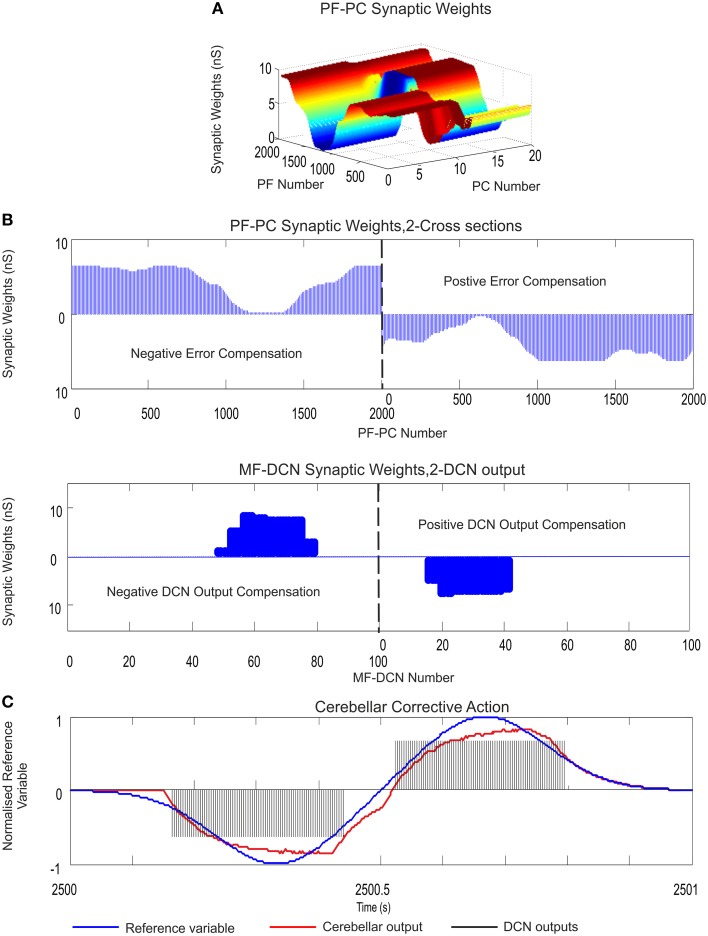
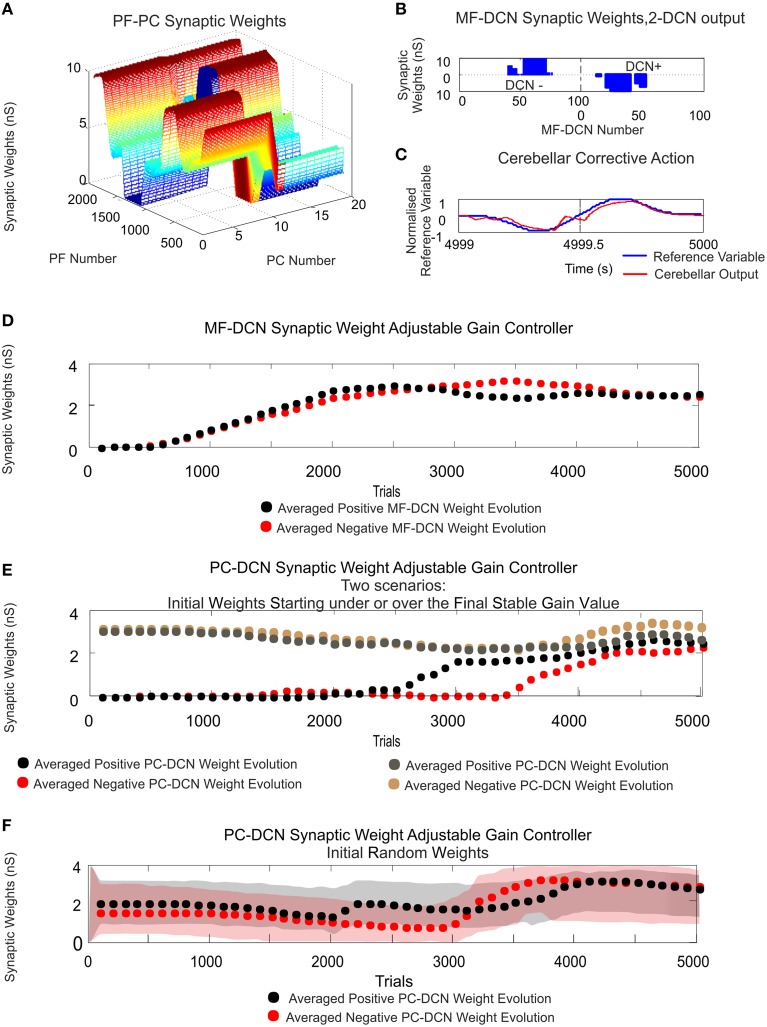
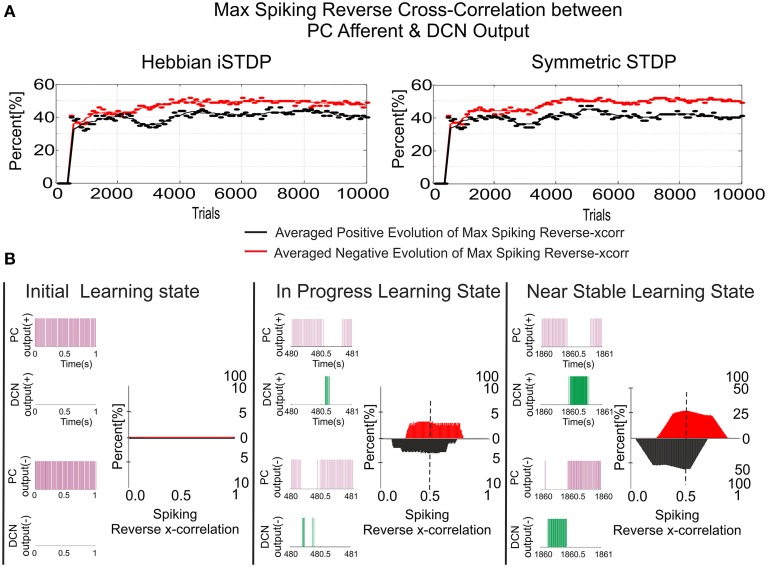
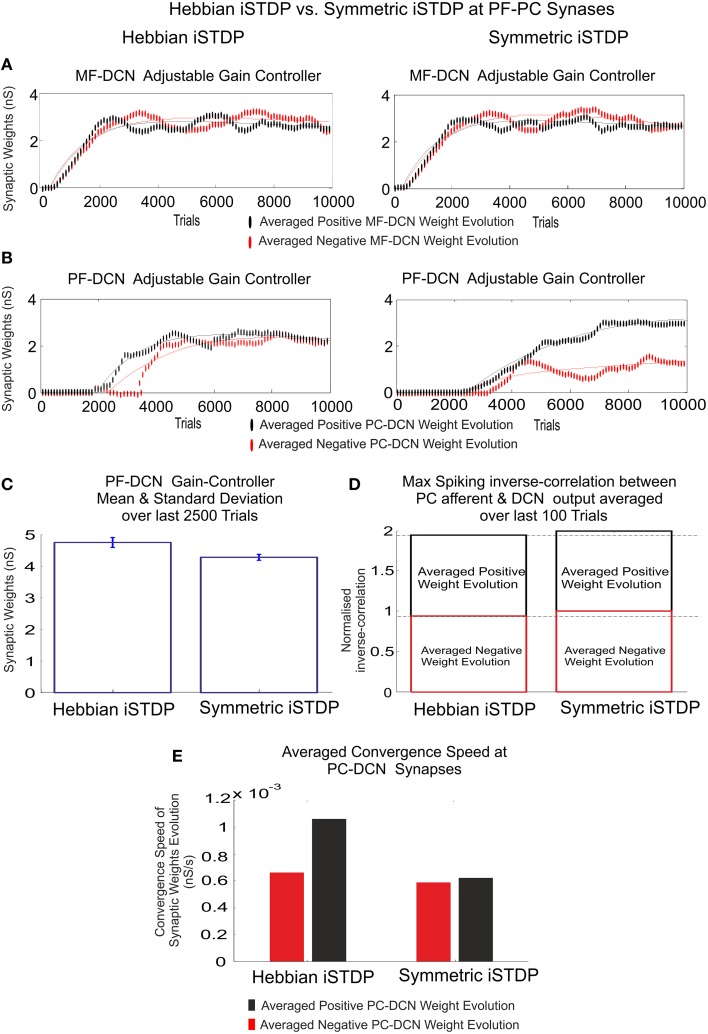
Deep cerebellar nuclei neurons receive both inhibitory (GABAergic) synaptic currents from Purkinje cells (within the cerebellar cortex) and excitatory (glutamatergic) synaptic currents from mossy fibers. Those two deep cerebellar nucleus inputs are thought to be also adaptive, embedding interesting properties in the framework of accurate movements. We show that distributed spike-timing-dependent plasticity mechanisms (STDP) located at different cerebellar sites (parallel fibers to Purkinje cells, mossy fibers to deep cerebellar nucleus cells, and Purkinje cells to deep cerebellar nucleus cells) in close-loop simulations provide an explanation for the complex learning properties of the cerebellum in motor learning. Concretely, we propose a new mechanistic cerebellar spiking model. In this new model, deep cerebellar nuclei embed a dual functionality: deep cerebellar nuclei acting as a gain adaptation mechanism and as a facilitator for the slow memory consolidation at mossy fibers to deep cerebellar nucleus synapses. Equipping the cerebellum with excitatory (e-STDP) and inhibitory (i-STDP) mechanisms at deep cerebellar nuclei afferents allows the accommodation of synaptic memories that were formed at parallel fibers to Purkinje cells synapses and then transferred to mossy fibers to deep cerebellar nucleus synapses. These adaptive mechanisms also contribute to modulate the deep-cerebellar-nucleus-output firing rate (output gain modulation toward optimizing its working range).
小脑深部核团神经元既接收来自(小脑皮质内的)浦肯野细胞的抑制性(γ-氨基丁酸能)突触电流,也接收来自苔藓纤维的兴奋性(谷氨酸能)突触电流。这两种小脑深部核团输入被认为也是适应性的,在精确运动的框架中嵌入了有趣的特性。我们表明,在闭环模拟中,位于不同小脑部位(平行纤维到浦肯野细胞、苔藓纤维到小脑深部核团细胞、浦肯野细胞到小脑深部核团细胞)的分布式尖峰时间依赖可塑性机制(STDP)为小脑在运动学习中的复杂学习特性提供了解释。具体而言,我们提出了一种新的小脑尖峰机制模型。在这个新模型中,小脑深部核团具有双重功能:小脑深部核团作为增益适应机制,并作为苔藓纤维到小脑深部核团突触处缓慢记忆巩固的促进者。在小脑深部核团传入纤维处配备兴奋性(e-STDP)和抑制性(i-STDP)机制,能够容纳在平行纤维到浦肯野细胞突触处形成、然后转移到苔藓纤维到小脑深部核团突触处的突触记忆。这些适应性机制也有助于调节小脑深部核团输出放电率(朝着优化其工作范围进行输出增益调节)。