Goryachev Andrew B, Leda Marcin, Miller Ann L, von Dassow George, Bement William M
a Centre for Systems and Synthetic Biology, Cell Biology Institute, School of Biological Sciences, University of Edinburgh , Edinburgh , UK.
b Department of Molecular , Cellular and Developmental Biology, University of Michigan , Ann-Arbor , MI , USA.
Small GTPases. 2016 Apr 2;7(2):65-70. doi: 10.1080/21541248.2016.1168505. Epub 2016 Apr 12.
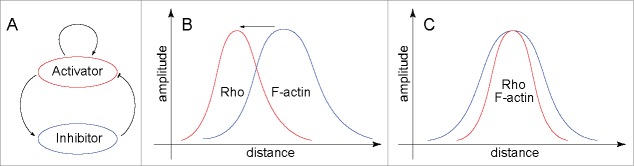
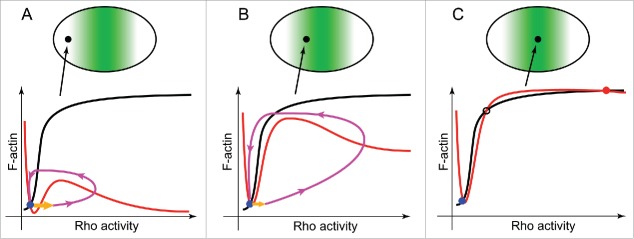
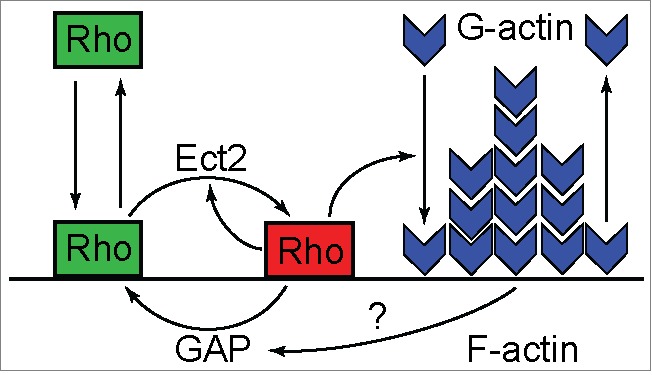
Emergence of the cytokinetic Rho zone that orchestrates formation and ingression of the cleavage furrow had been explained previously via microtubule-dependent cortical concentration of Ect2, a guanine nucleotide exchange factor for Rho. The results of a recent publication now demonstrate that, en route from resting cortex to fully established furrow, there lies a regime of cortical excitability in which Rho activity and F-actin play the roles of the prototypical activator and inhibitor, respectively. This cortical excitability is manifest as dramatic traveling waves on the cortex of oocytes and embryos of frogs and starfish. These waves are initiated by autocatalytic activation of Rho at the wave front and extinguished by F-actin-dependent inhibition at their back. It is still unclear how propagating excitable Rho-actin waves give rise to the stable co-existence of Rho activity and F-actin density in the static cleavage furrow during cytokinesis. It is possible that some central spindle-associated signaling molecule simply turns off the inhibition of Rho activity by F-actin. However, mathematical modeling suggests a distinct scenario in which local "re-wiring" of the Rho-actin coupling in the furrow is no longer necessary. Instead, the model predicts that the continuously rising level of Ect2 produces in the furrow a qualitatively new stable steady state that replaces excitability and brings about the stable co-existence of high Rho activity and dense F-actin despite the continuing inhibition of Rho by F-actin.
之前曾通过微管依赖性的Ect2(一种Rho的鸟嘌呤核苷酸交换因子)在皮质的聚集来解释协调分裂沟形成和内陷的细胞动力学Rho区的出现。最近一篇发表文章的结果现在表明,在从静止皮质到完全形成的分裂沟的过程中,存在一种皮质兴奋性状态,其中Rho活性和F-肌动蛋白分别起着典型激活剂和抑制剂的作用。这种皮质兴奋性在青蛙和海星的卵母细胞和胚胎的皮质上表现为显著的行波。这些波由波前的Rho自催化激活引发,并在波后由F-肌动蛋白依赖性抑制而熄灭。目前仍不清楚在胞质分裂过程中,传播的可兴奋Rho-肌动蛋白波如何在静态分裂沟中导致Rho活性和F-肌动蛋白密度的稳定共存。有可能某些中央纺锤体相关信号分子只是简单地关闭了F-肌动蛋白对Rho活性的抑制。然而,数学模型提出了一种不同的情况,即不再需要在分裂沟中对Rho-肌动蛋白偶联进行局部“重新布线”。相反,该模型预测,Ect2水平的持续上升在分裂沟中产生了一种质的新稳定稳态,它取代了兴奋性,并导致尽管F-肌动蛋白持续抑制Rho,但仍有高Rho活性和密集F-肌动蛋白的稳定共存。