Wibowo Anjar, Becker Claude, Marconi Gianpiero, Durr Julius, Price Jonathan, Hagmann Jorg, Papareddy Ranjith, Putra Hadi, Kageyama Jorge, Becker Jorg, Weigel Detlef, Gutierrez-Marcos Jose
School of Life Sciences, University of Warwick, Coventry, United Kingdom.
Department of Molecular Biology, Max Planck Institute for Developmental Biology, Tübingen, Germany.
Elife. 2016 May 31;5:e13546. doi: 10.7554/eLife.13546.
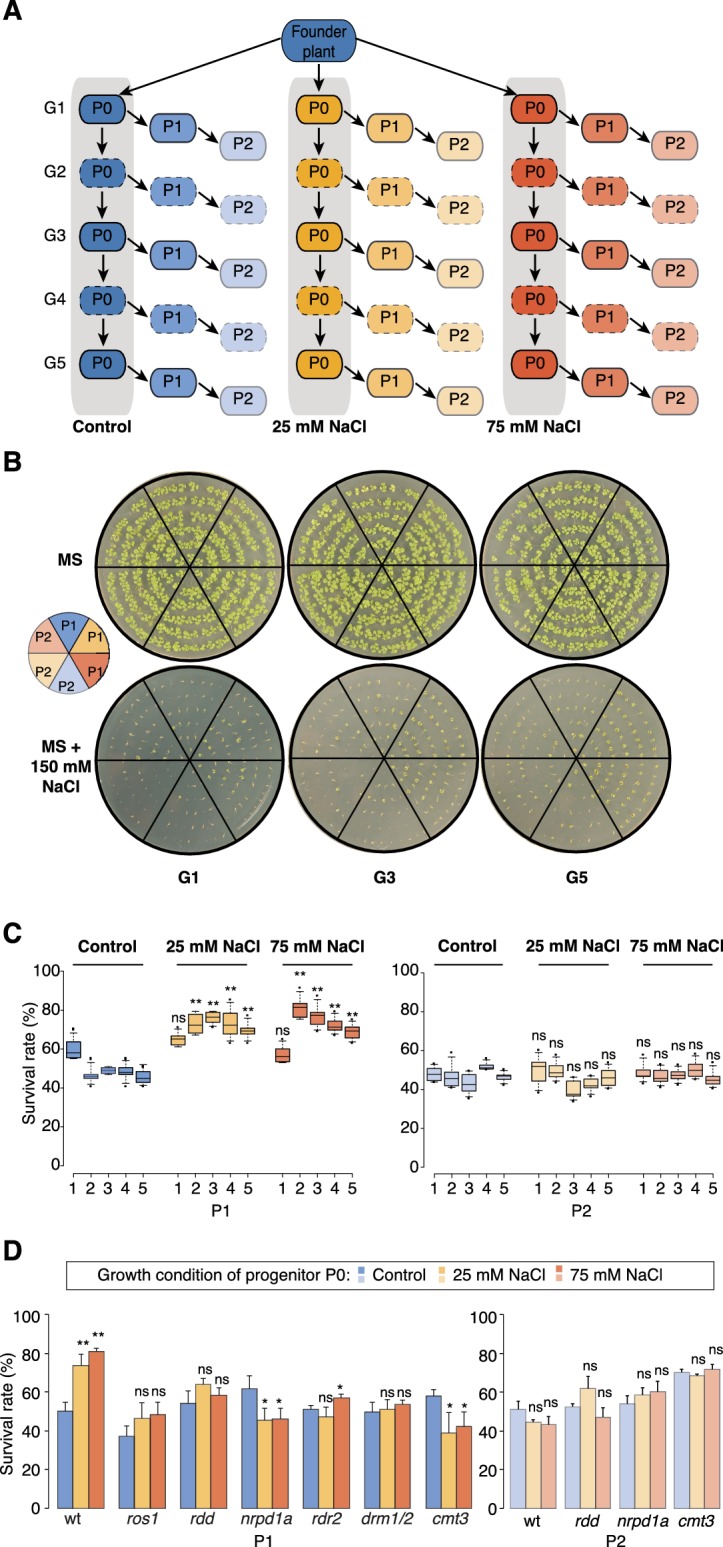
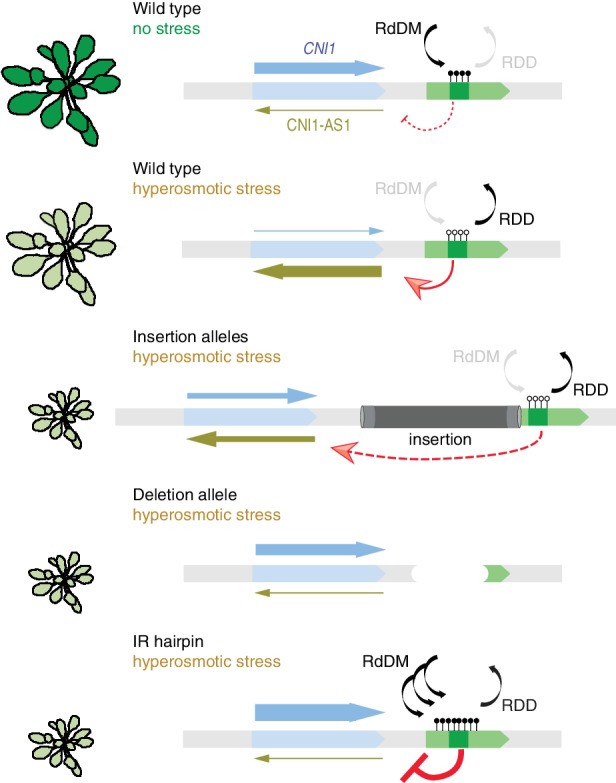
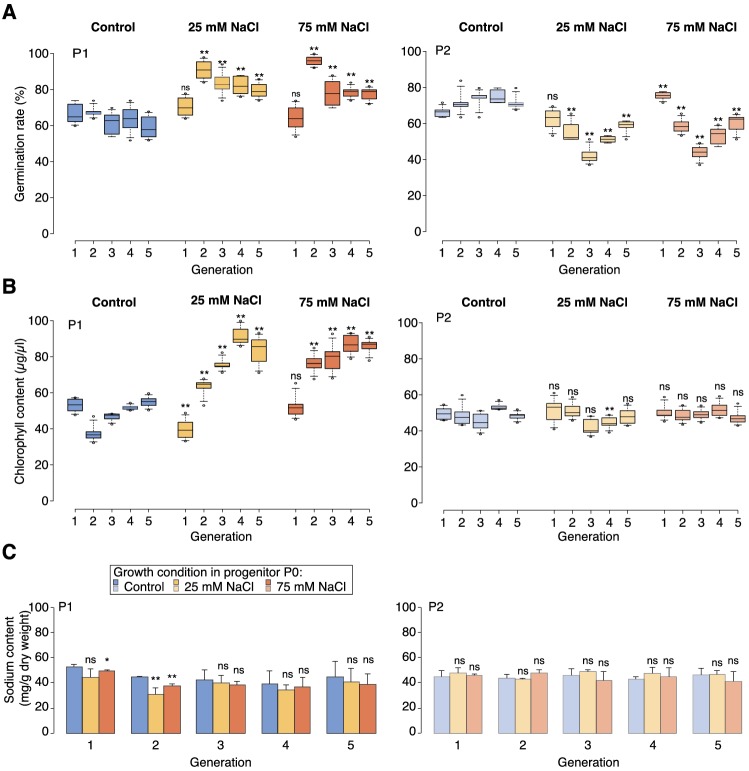
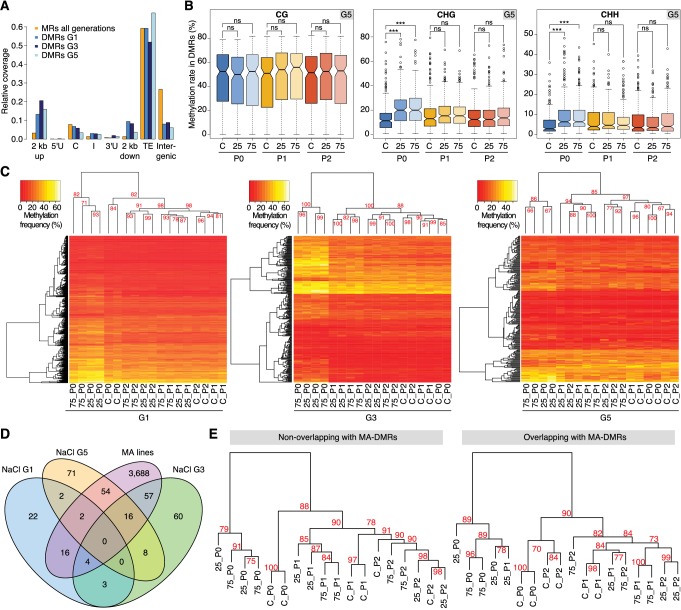
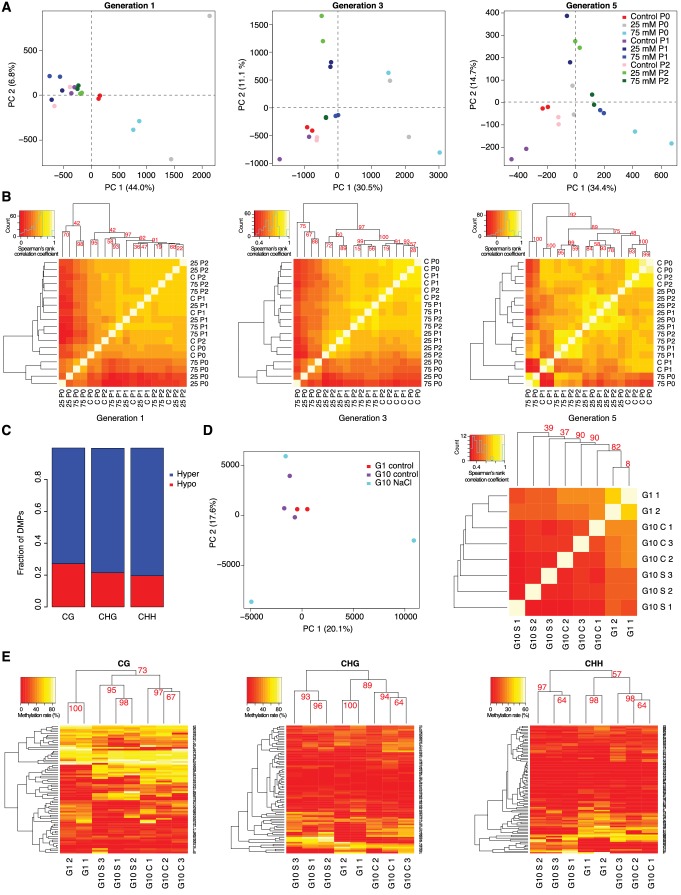
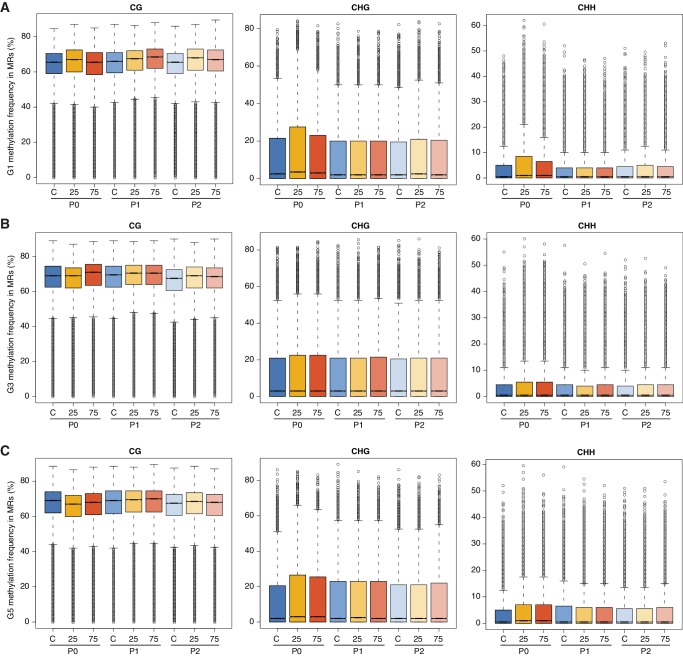
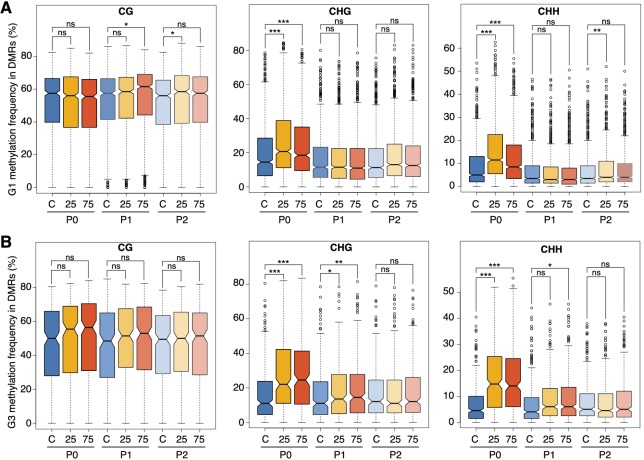
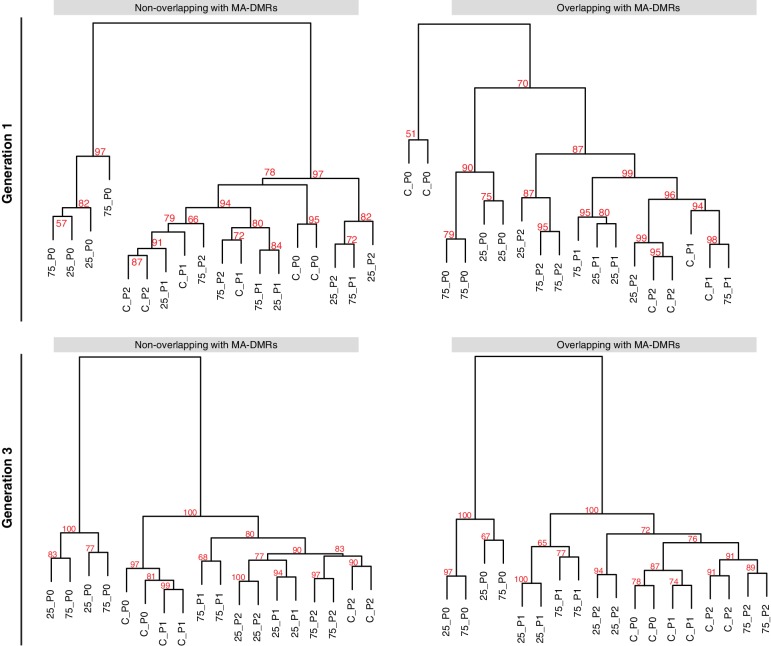
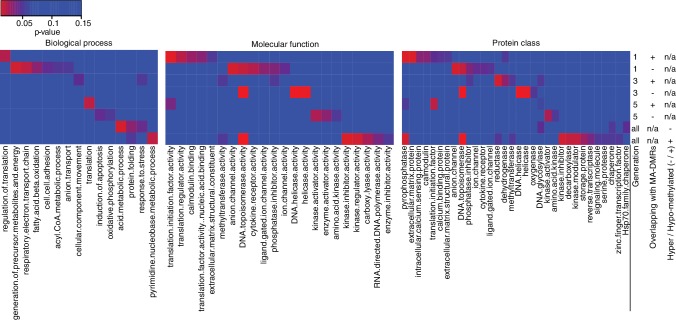
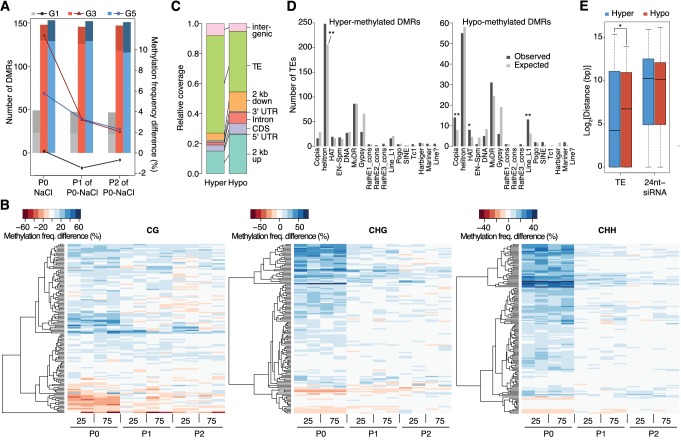
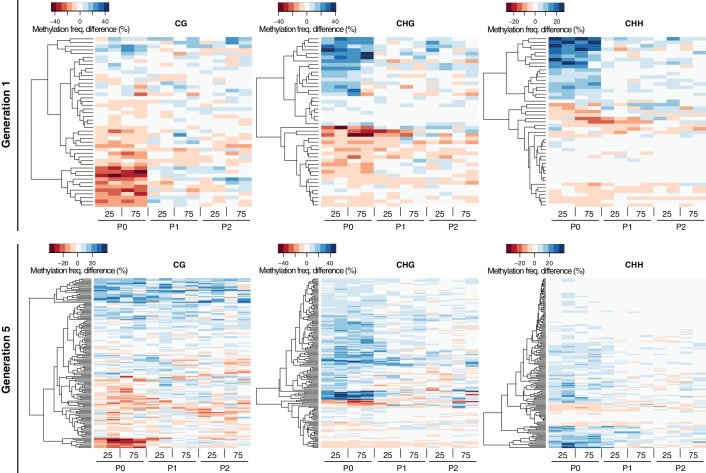
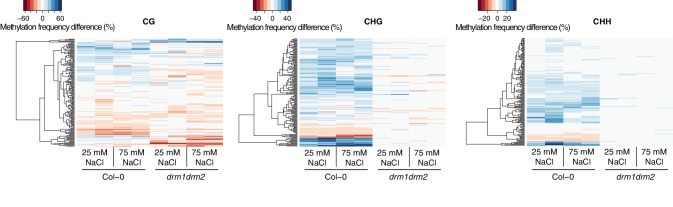
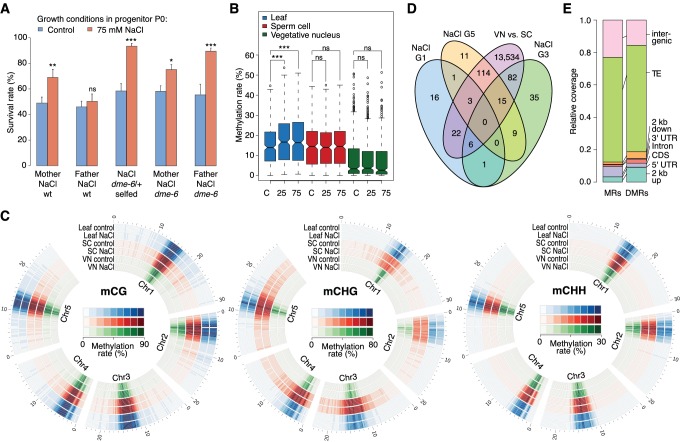
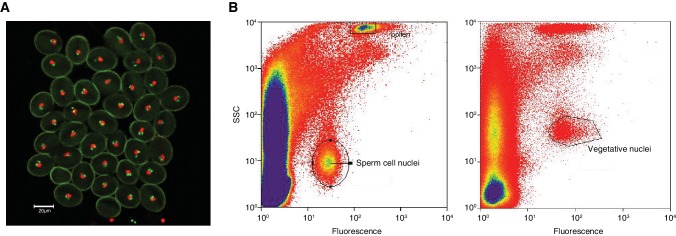
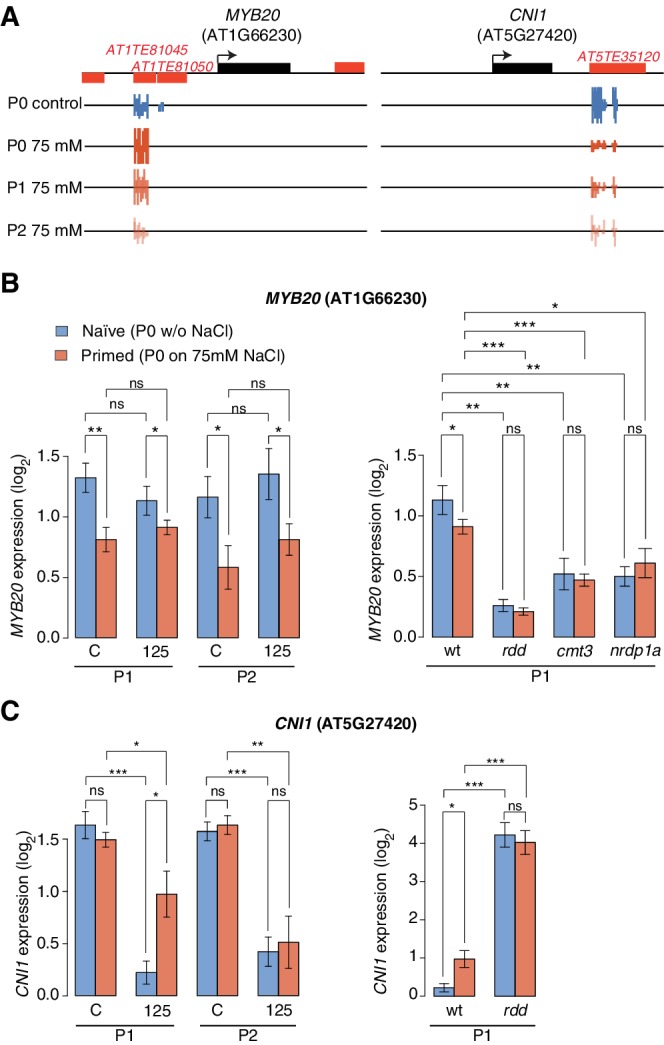
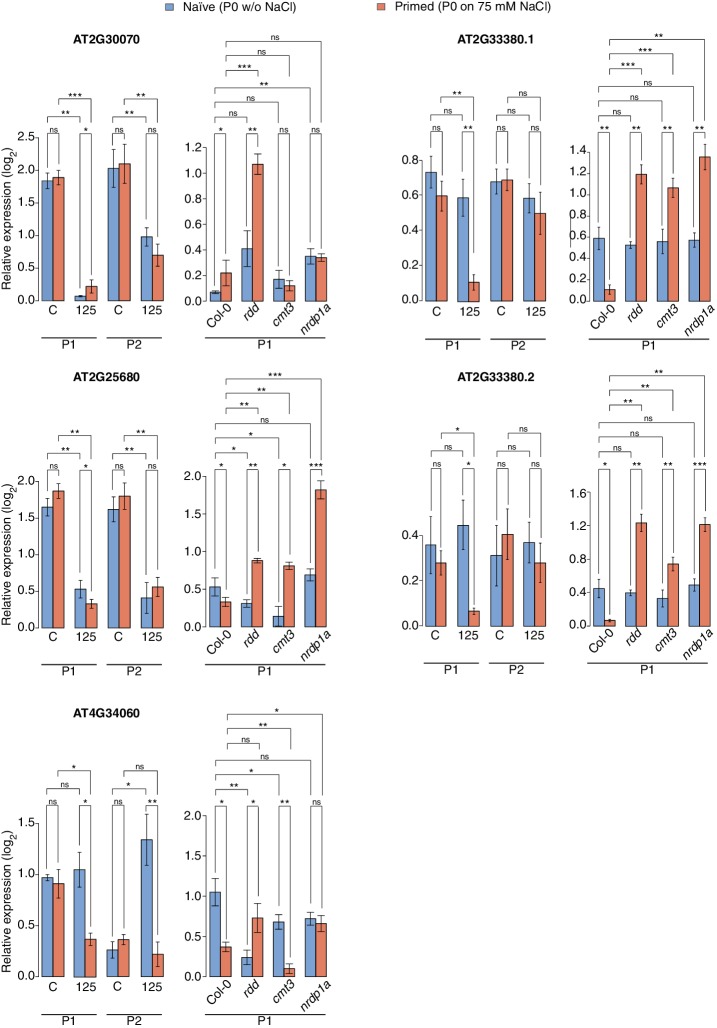
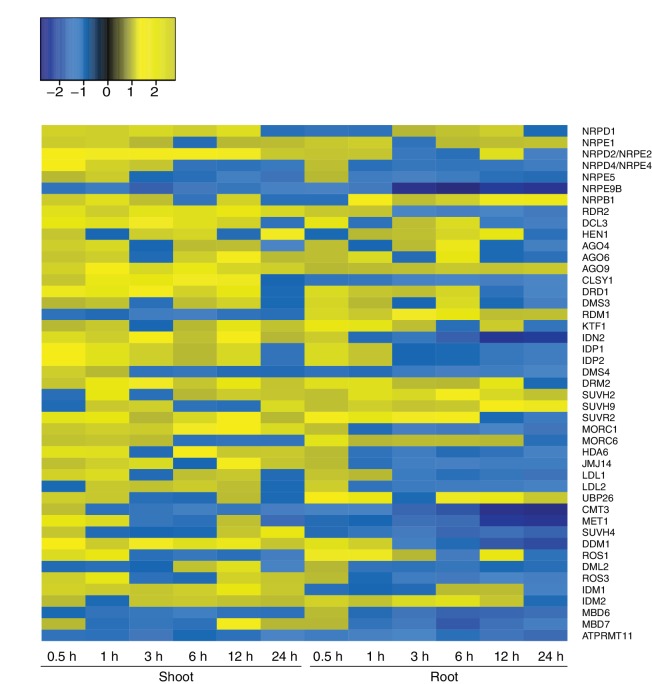
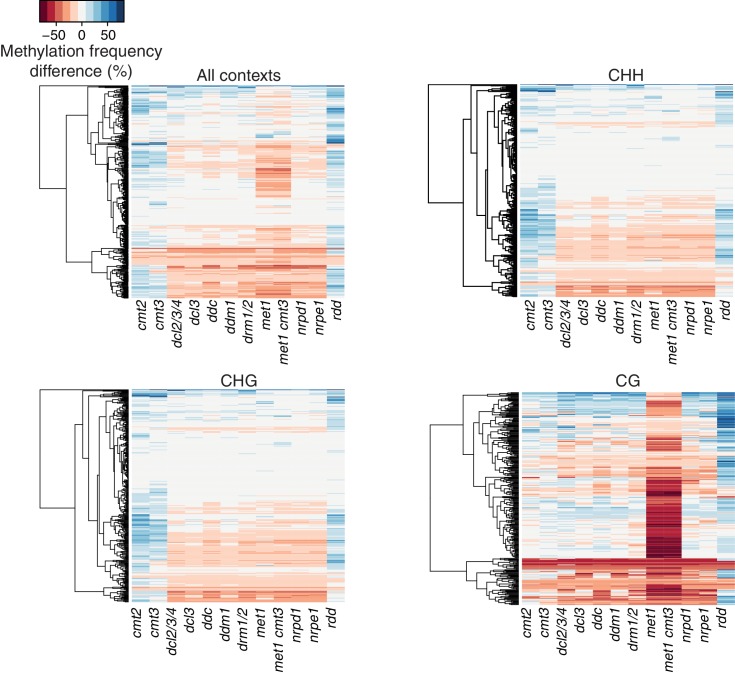
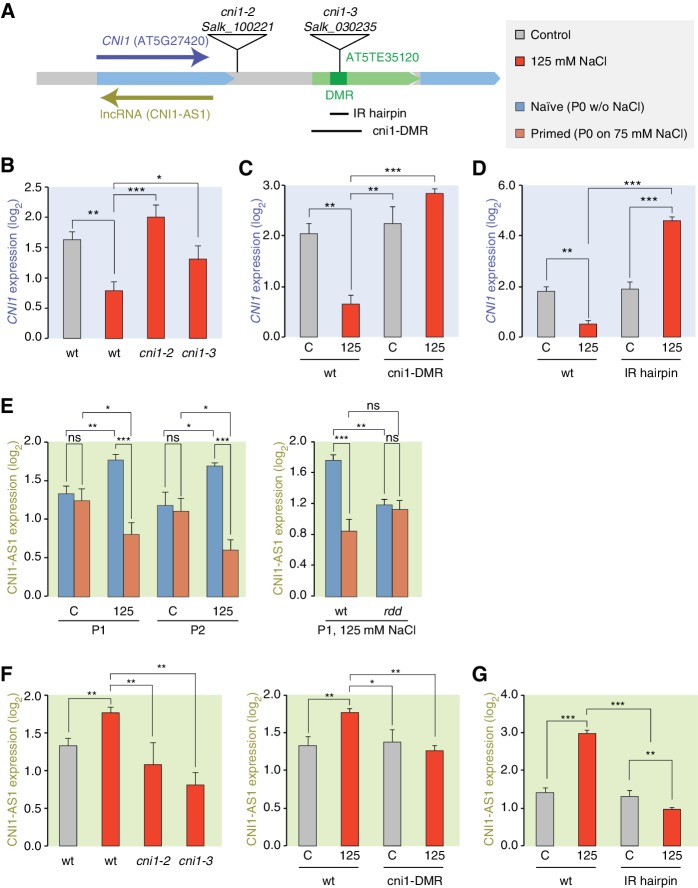
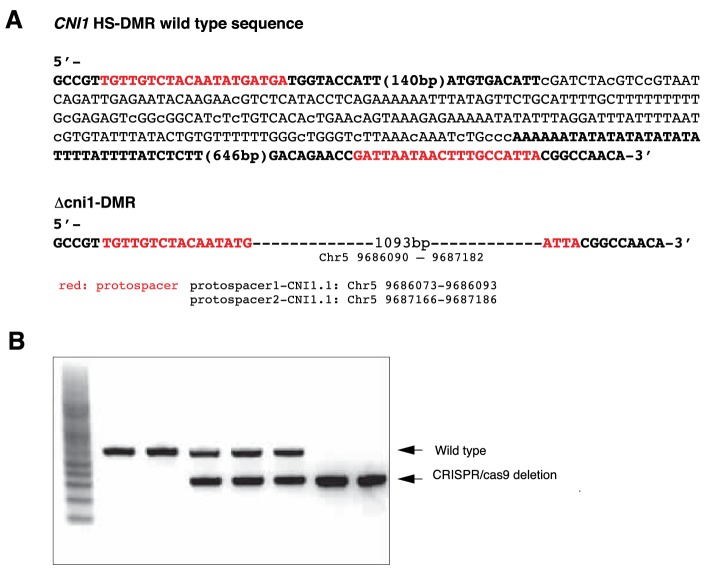
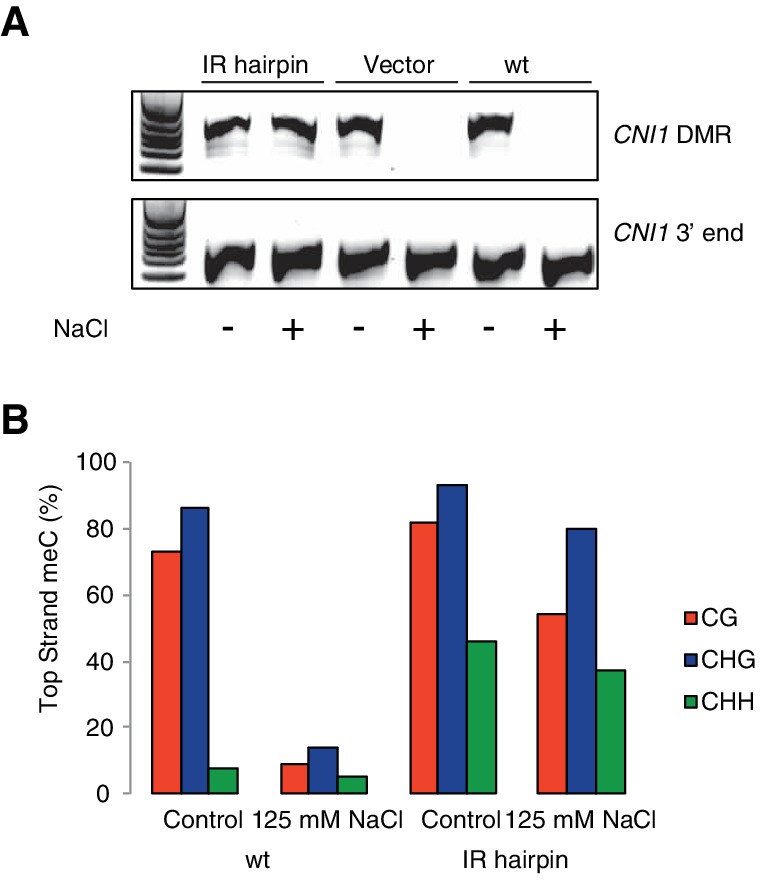
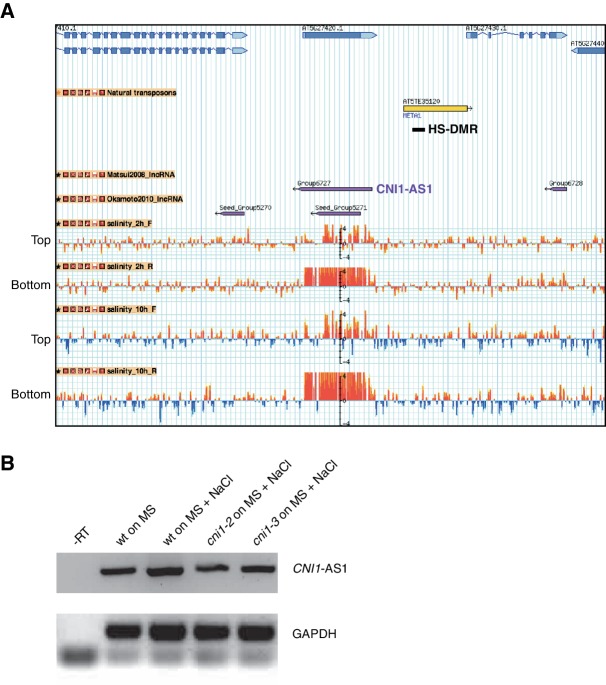
Inducible epigenetic changes in eukaryotes are believed to enable rapid adaptation to environmental fluctuations. We have found distinct regions of the Arabidopsis genome that are susceptible to DNA (de)methylation in response to hyperosmotic stress. The stress-induced epigenetic changes are associated with conditionally heritable adaptive phenotypic stress responses. However, these stress responses are primarily transmitted to the next generation through the female lineage due to widespread DNA glycosylase activity in the male germline, and extensively reset in the absence of stress. Using the CNI1/ATL31 locus as an example, we demonstrate that epigenetically targeted sequences function as distantly-acting control elements of antisense long non-coding RNAs, which in turn regulate targeted gene expression in response to stress. Collectively, our findings reveal that plants use a highly dynamic maternal 'short-term stress memory' with which to respond to adverse external conditions. This transient memory relies on the DNA methylation machinery and associated transcriptional changes to extend the phenotypic plasticity accessible to the immediate offspring.
真核生物中可诱导的表观遗传变化被认为能够使其快速适应环境波动。我们发现拟南芥基因组的不同区域在高渗胁迫下易发生DNA(去)甲基化。胁迫诱导的表观遗传变化与条件可遗传的适应性表型胁迫反应相关。然而,由于雄性生殖系中广泛存在的DNA糖基化酶活性,这些胁迫反应主要通过雌性谱系传递给下一代,并且在没有胁迫的情况下会大量重置。以CNI1/ATL31基因座为例,我们证明表观遗传靶向序列作为反义长链非编码RNA的远距离作用控制元件,进而响应胁迫调节靶向基因表达。总体而言,我们的研究结果表明植物利用高度动态的母本“短期胁迫记忆”来应对不利的外部条件。这种短暂的记忆依赖于DNA甲基化机制和相关的转录变化,以扩展直接后代可获得的表型可塑性。