Whitelaw Jamie A, Latorre-Barragan Fernanda, Gras Simon, Pall Gurman S, Leung Jacqueline M, Heaslip Aoife, Egarter Saskia, Andenmatten Nicole, Nelson Shane R, Warshaw David M, Ward Gary E, Meissner Markus
Wellcome Trust Centre For Molecular Parasitology, Institute of Infection, Immunity & Inflammation, Glasgow Biomedical Research Centre, University of Glasgow, 120 University Place, Glasgow, G12 8TA, UK.
Department of Biology, Indiana University, Bloomington, Myers Hall 240, 915 E 3rd St Bloomington, Bloomington, IN, 47405, USA.
BMC Biol. 2017 Jan 18;15(1):1. doi: 10.1186/s12915-016-0343-5.
Apicomplexan parasites employ a unique form of movement, termed gliding motility, in order to invade the host cell. This movement depends on the parasite's actomyosin system, which is thought to generate the force during gliding. However, recent evidence questions the exact molecular role of this system, since mutants for core components of the gliding machinery, such as parasite actin or subunits of the MyoA-motor complex (the glideosome), remain motile and invasive, albeit at significantly reduced efficiencies. While compensatory mechanisms and unusual polymerisation kinetics of parasite actin have been evoked to explain these findings, the actomyosin system could also play a role distinct from force production during parasite movement.
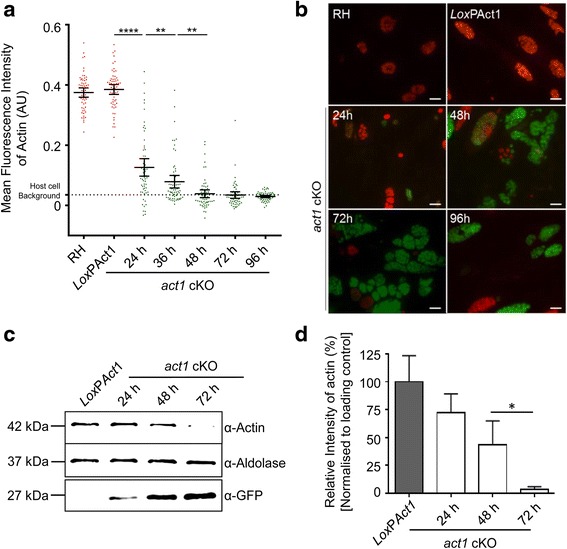
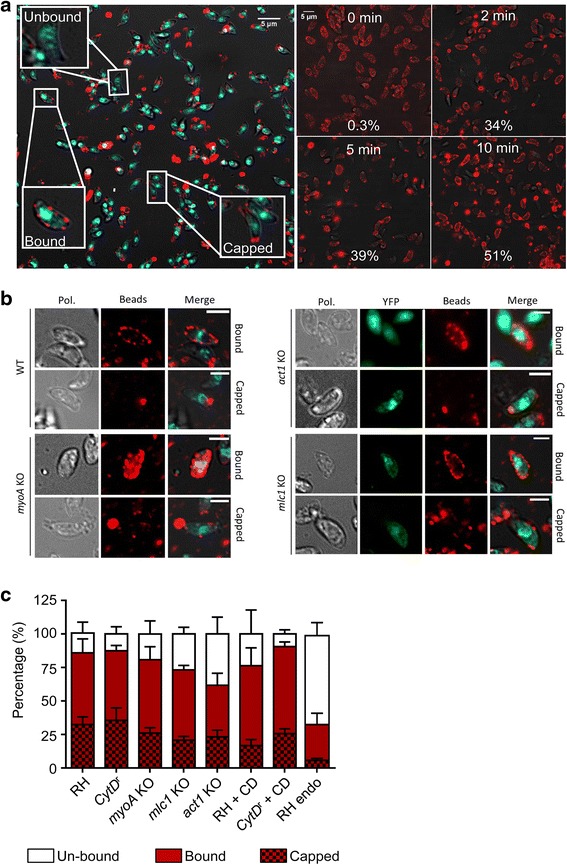
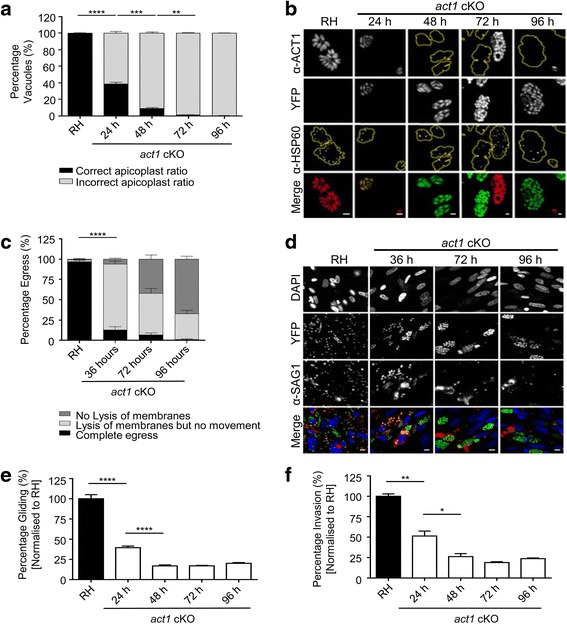
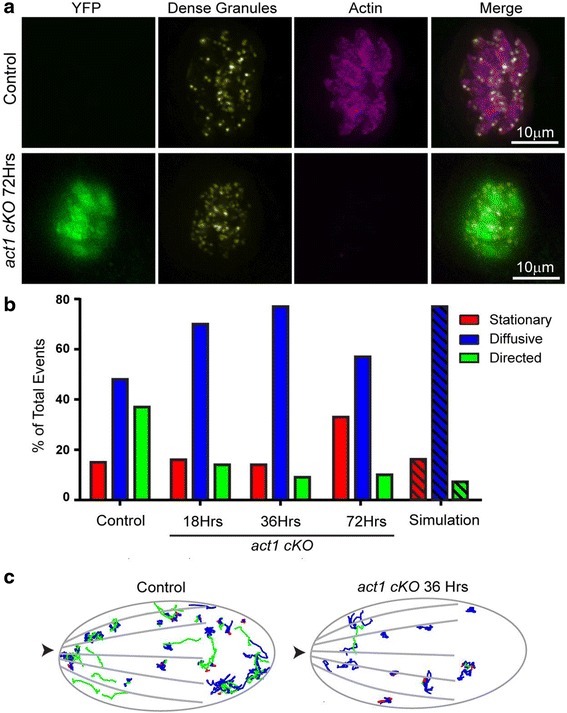
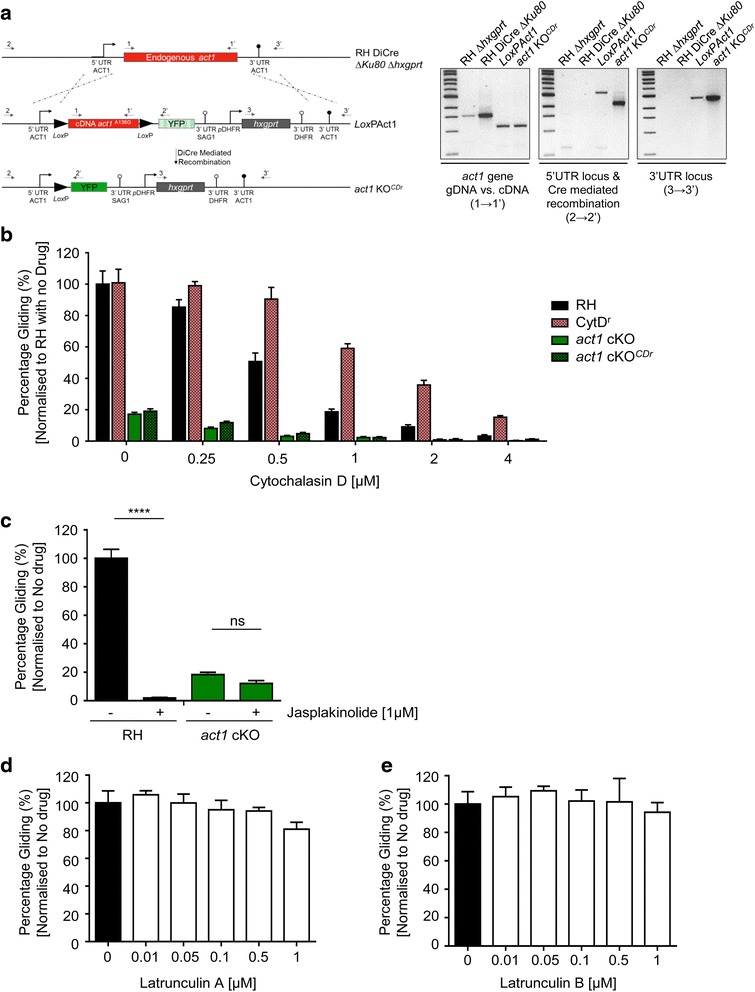
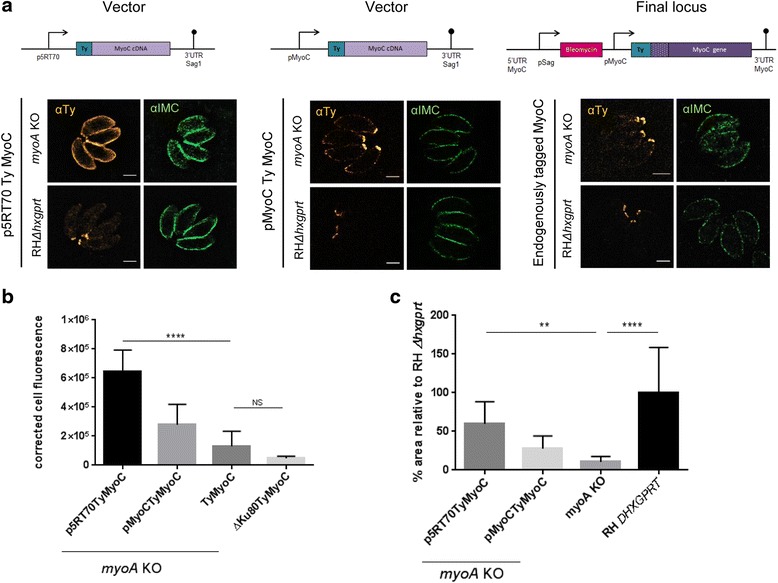
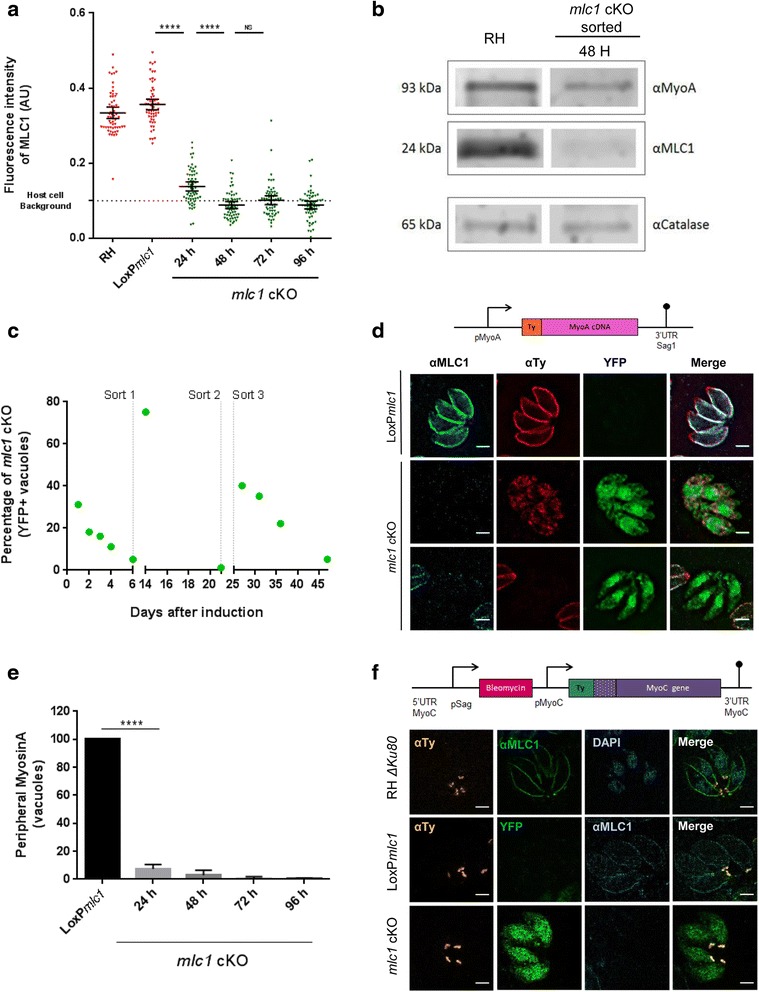
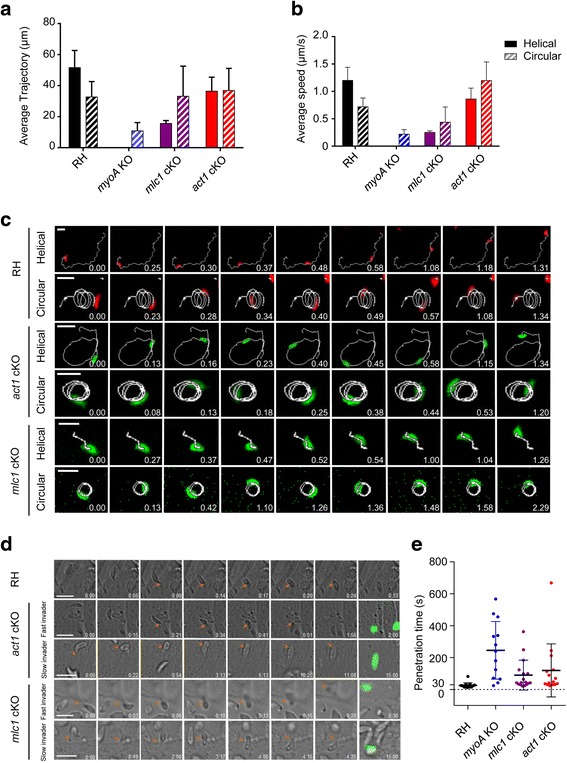
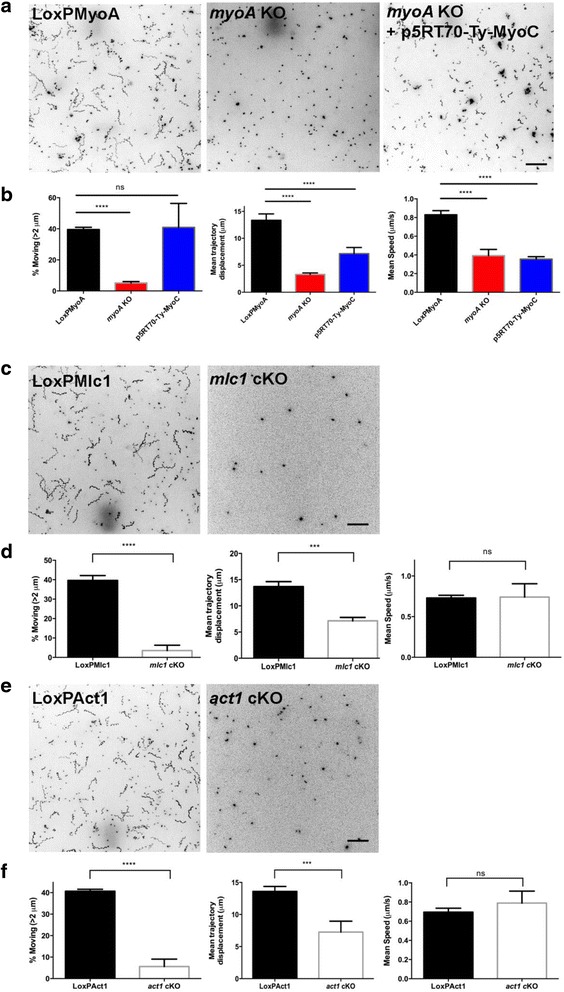
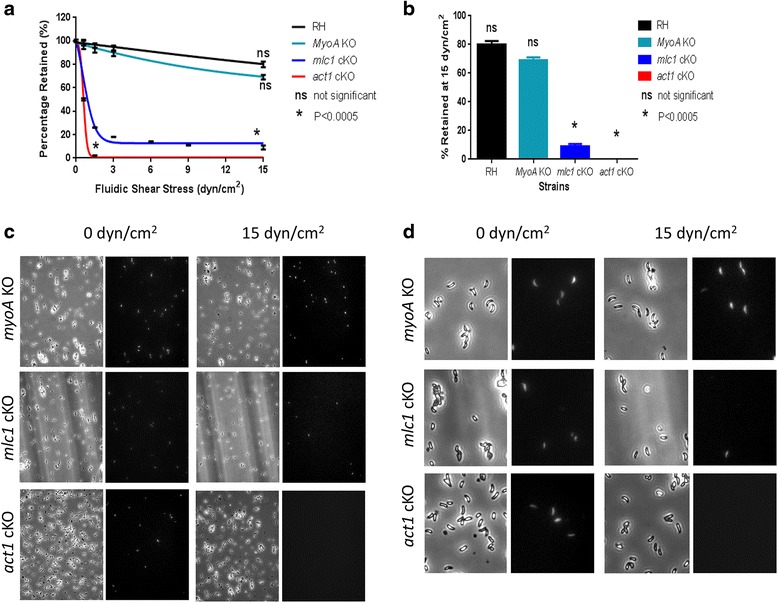
In this study, we compared the phenotypes of different mutants for core components of the actomyosin system in Toxoplasma gondii to decipher their exact role during gliding motility and invasion. We found that, while some phenotypes (apicoplast segregation, host cell egress, dense granule motility) appeared early after induction of the act1 knockout and went to completion, a small percentage of the parasites remained capable of motility and invasion well past the point at which actin levels were undetectable. Those act1 conditional knockout (cKO) and mlc1 cKO that continue to move in 3D do so at speeds similar to wildtype parasites. However, these mutants are virtually unable to attach to a collagen-coated substrate under flow conditions, indicating an important role for the actomyosin system of T. gondii in the formation of attachment sites.
We demonstrate that parasite actin is essential during the lytic cycle and cannot be compensated by other molecules. Our data suggest a conventional polymerisation mechanism in vivo that depends on a critical concentration of G-actin. Importantly, we demonstrate that the actomyosin system of the parasite functions in attachment to the surface substrate, and not necessarily as force generator.
顶复门寄生虫采用一种独特的运动形式,即滑行运动,以便侵入宿主细胞。这种运动依赖于寄生虫的肌动球蛋白系统,该系统被认为在滑行过程中产生力量。然而,最近的证据对该系统的确切分子作用提出了质疑,因为滑行机制核心成分的突变体,如寄生虫肌动蛋白或肌动蛋白A运动复合体(滑行体)的亚基,尽管效率显著降低,但仍具有运动性和侵袭性。虽然已经提出了寄生虫肌动蛋白的补偿机制和异常聚合动力学来解释这些发现,但肌动球蛋白系统在寄生虫运动过程中也可能发挥与力量产生不同的作用。
在本研究中,我们比较了刚地弓形虫肌动球蛋白系统核心成分不同突变体的表型,以解读它们在滑行运动和侵袭过程中的确切作用。我们发现,虽然一些表型(质体分离、宿主细胞逸出、致密颗粒运动)在诱导肌动蛋白1基因敲除后早期出现并完成,但一小部分寄生虫在肌动蛋白水平无法检测到之后很长时间仍具有运动性和侵袭性。那些在三维空间中继续移动的肌动蛋白1条件性基因敲除(cKO)和肌球蛋白轻链1 cKO寄生虫的移动速度与野生型寄生虫相似。然而,这些突变体在流动条件下几乎无法附着在胶原包被的底物上,这表明刚地弓形虫的肌动球蛋白系统在附着位点形成中起重要作用。
我们证明寄生虫肌动蛋白在裂解周期中是必不可少的,不能被其他分子补偿。我们的数据表明体内存在一种依赖于G-肌动蛋白临界浓度的传统聚合机制。重要的是,我们证明寄生虫的肌动球蛋白系统在附着于表面底物时起作用,而不一定作为力量产生器。