van Pee Katharina, Neuhaus Alexander, D'Imprima Edoardo, Mills Deryck J, Kühlbrandt Werner, Yildiz Özkan
Department of Structural Biology, Max Planck Institute of Biophysics, Frankfurt am Main, Germany.
Elife. 2017 Mar 21;6:e23644. doi: 10.7554/eLife.23644.
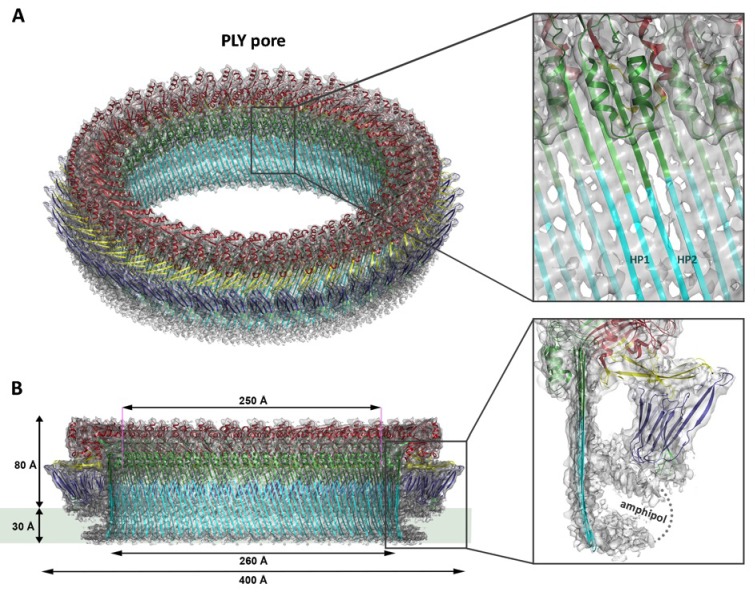
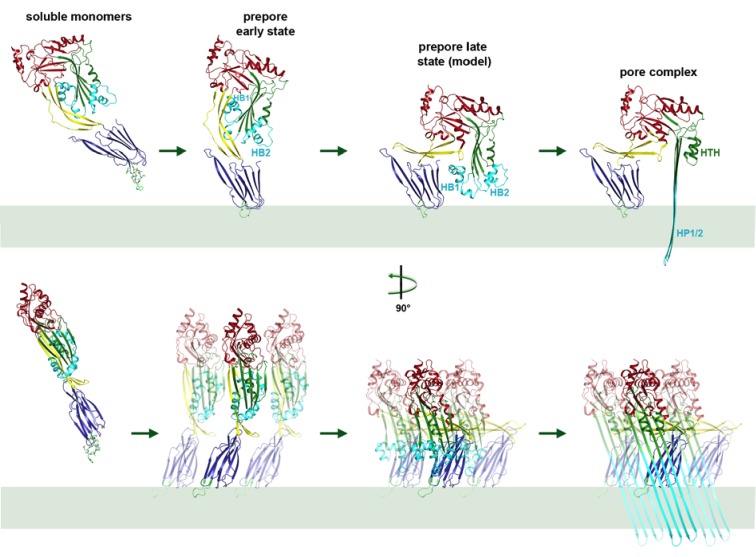
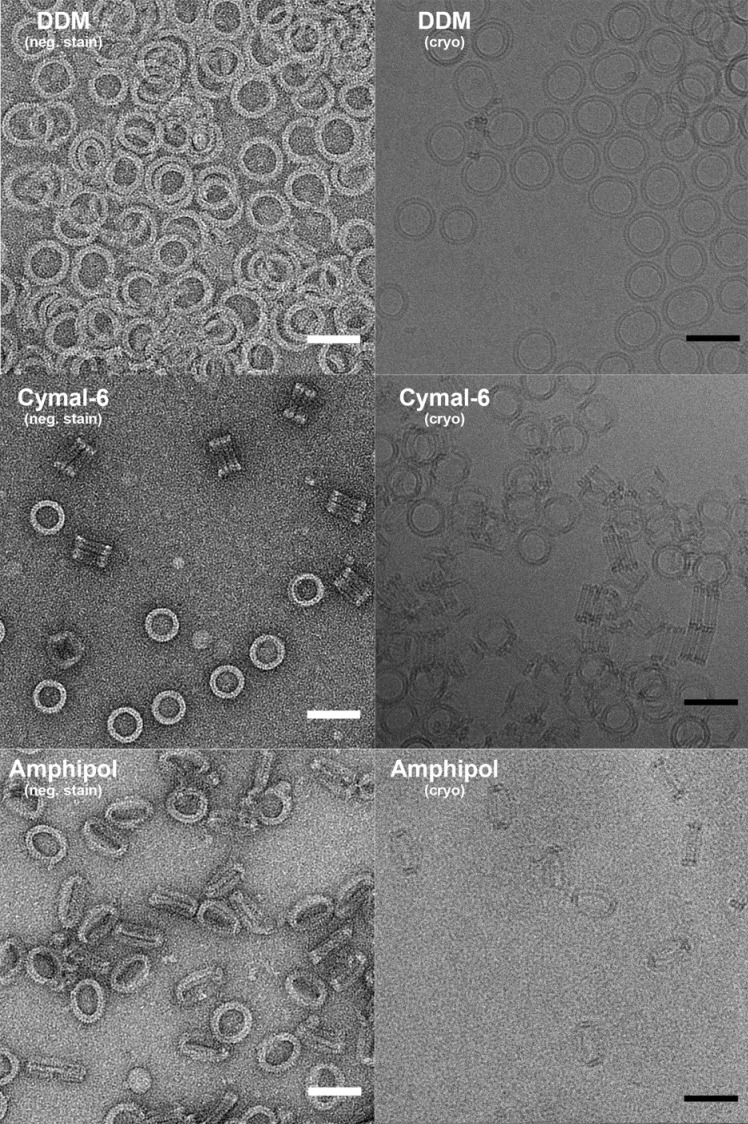
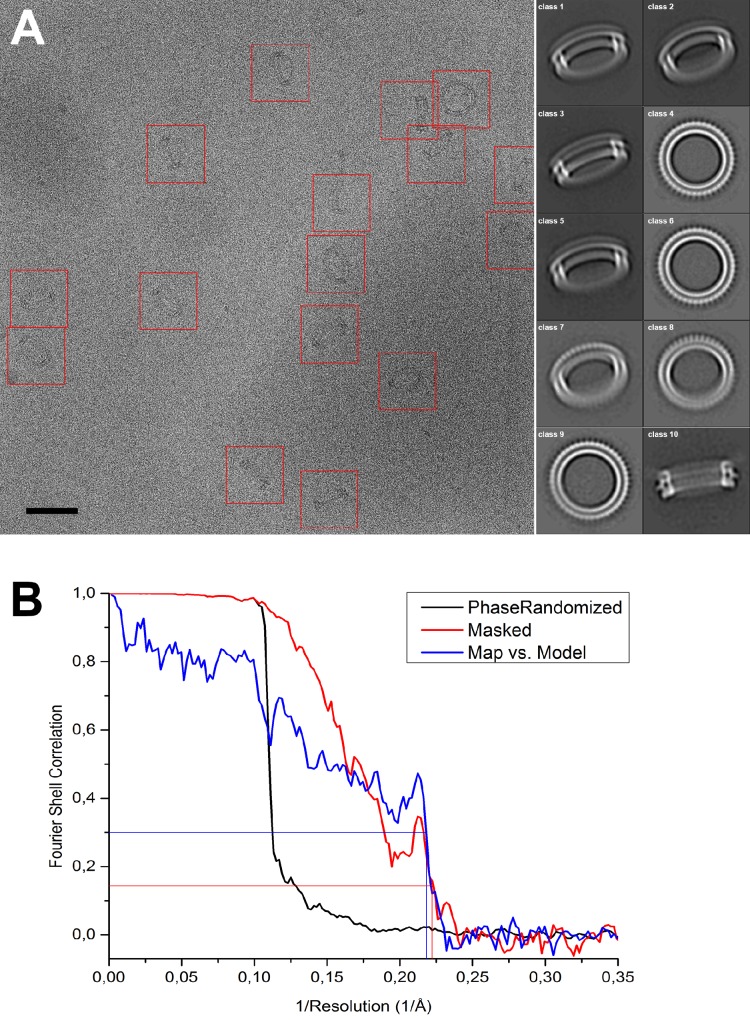
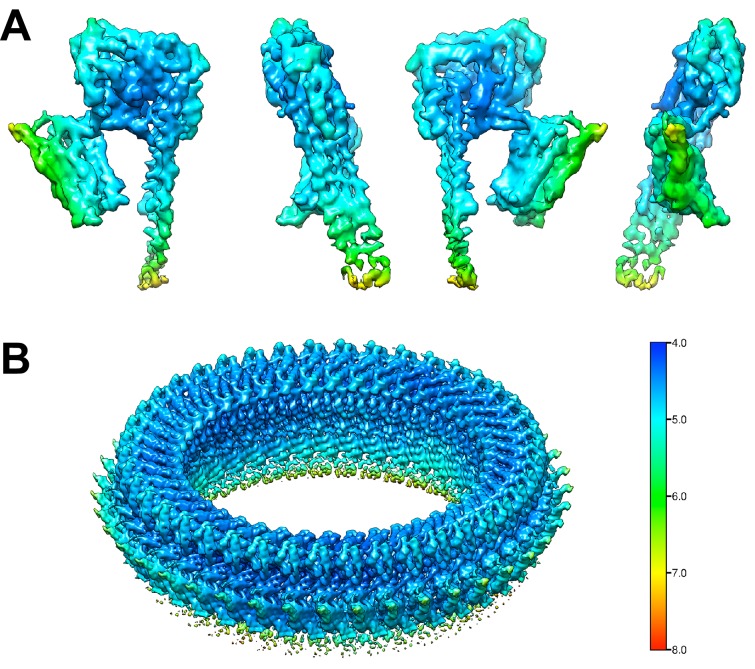
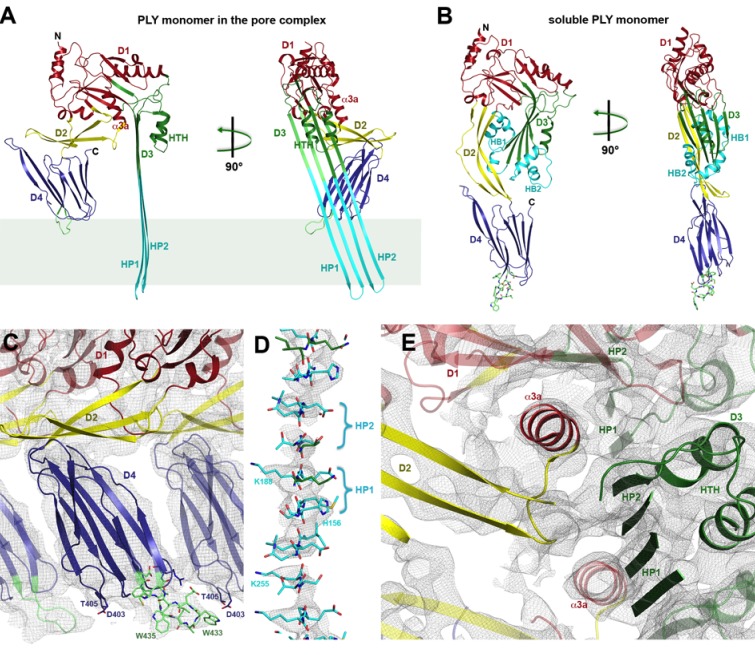
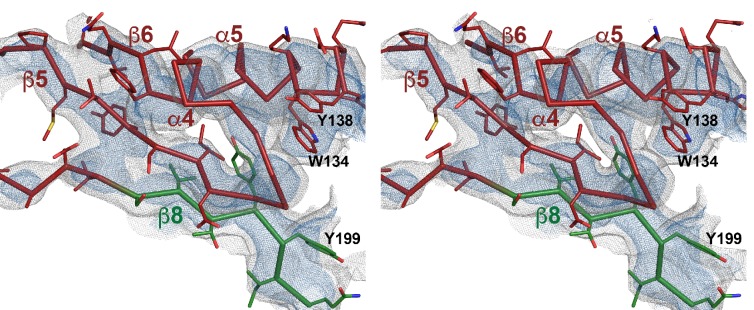
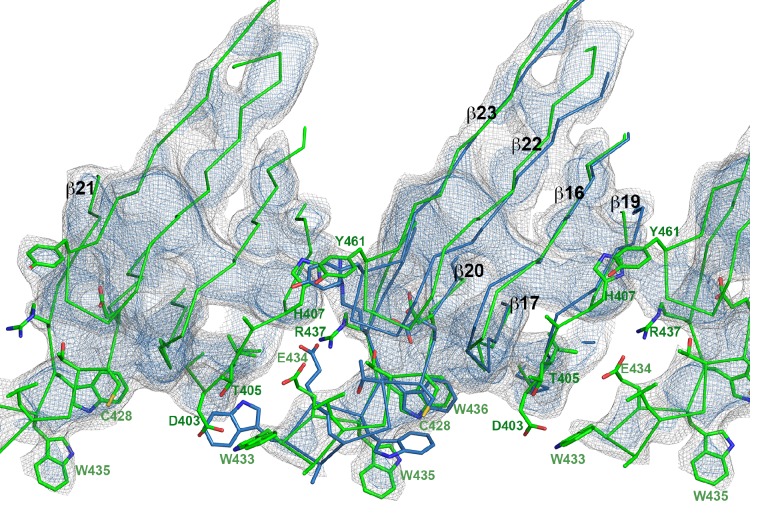
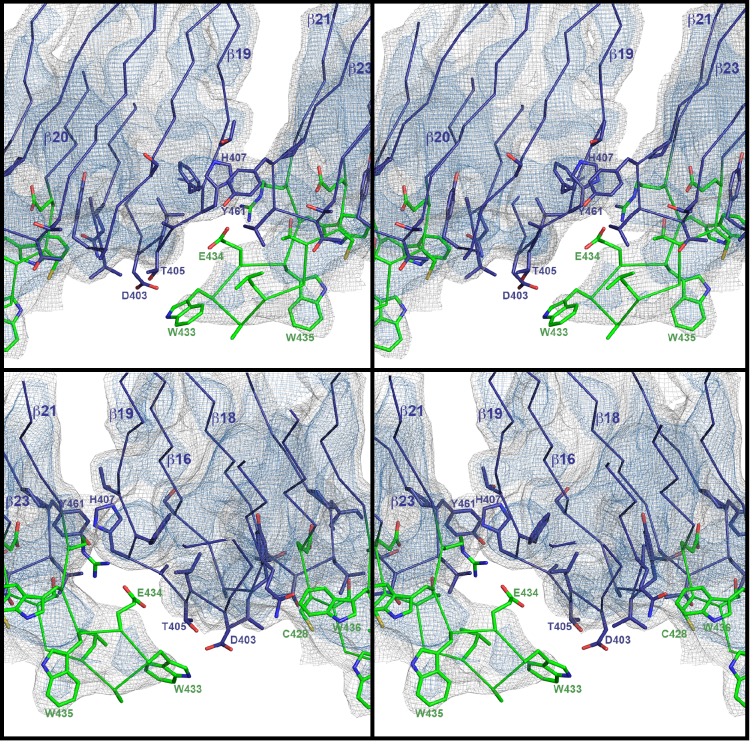
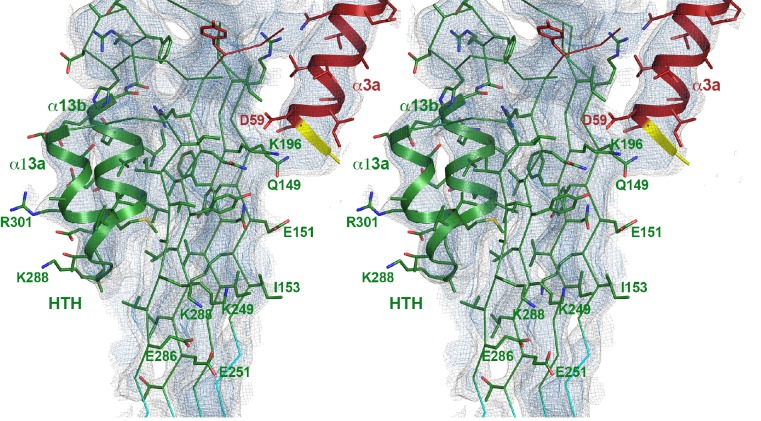
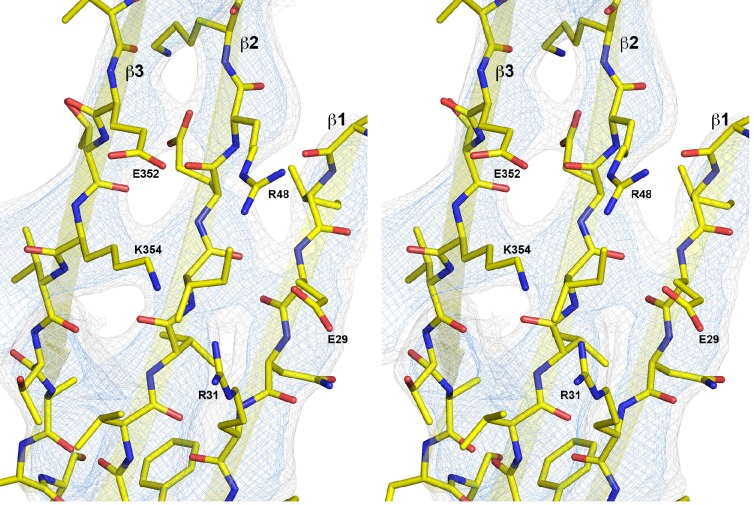
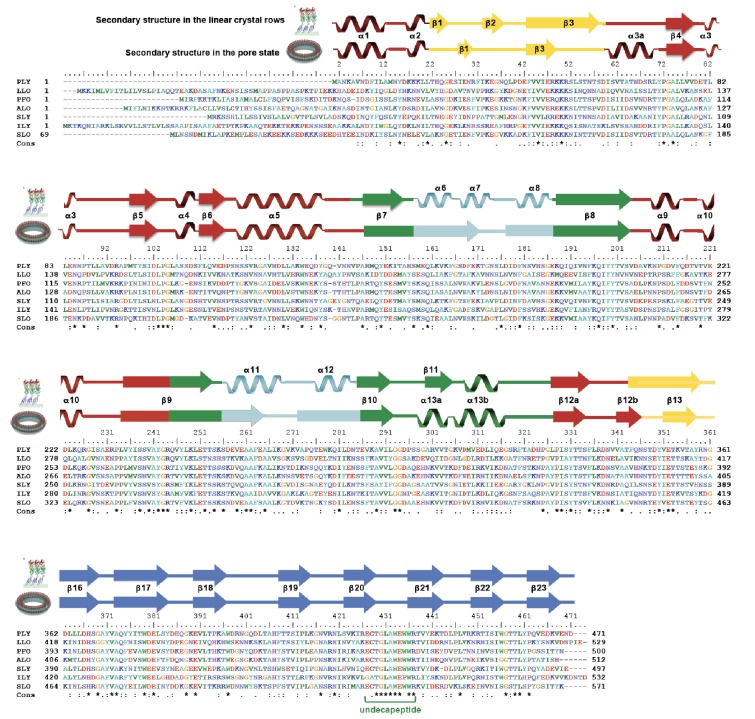
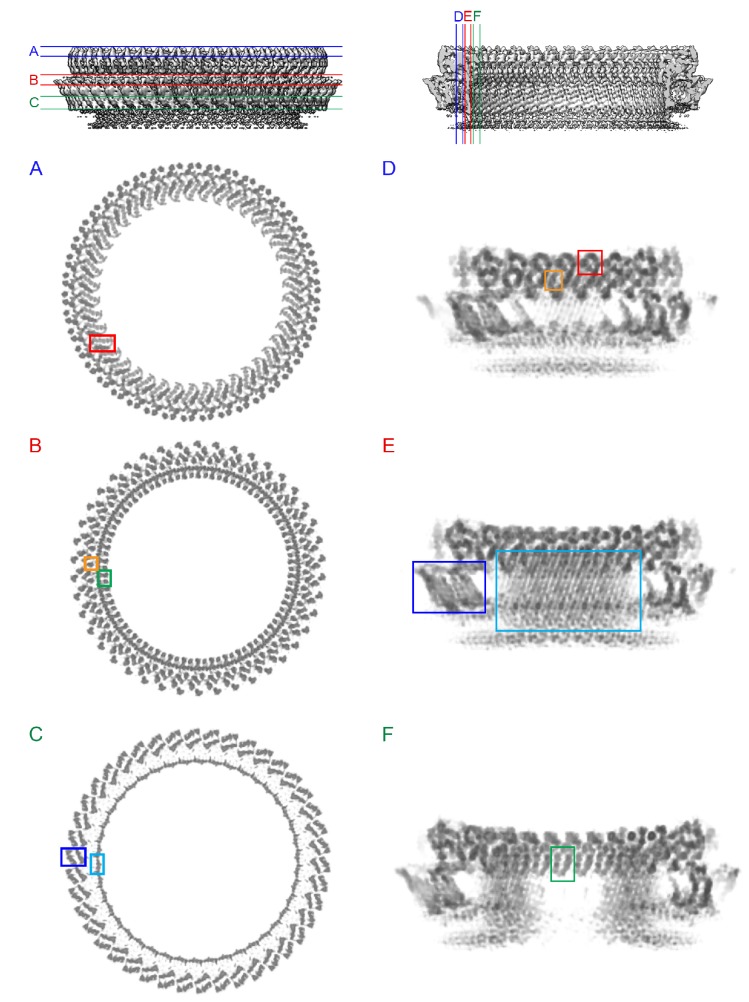
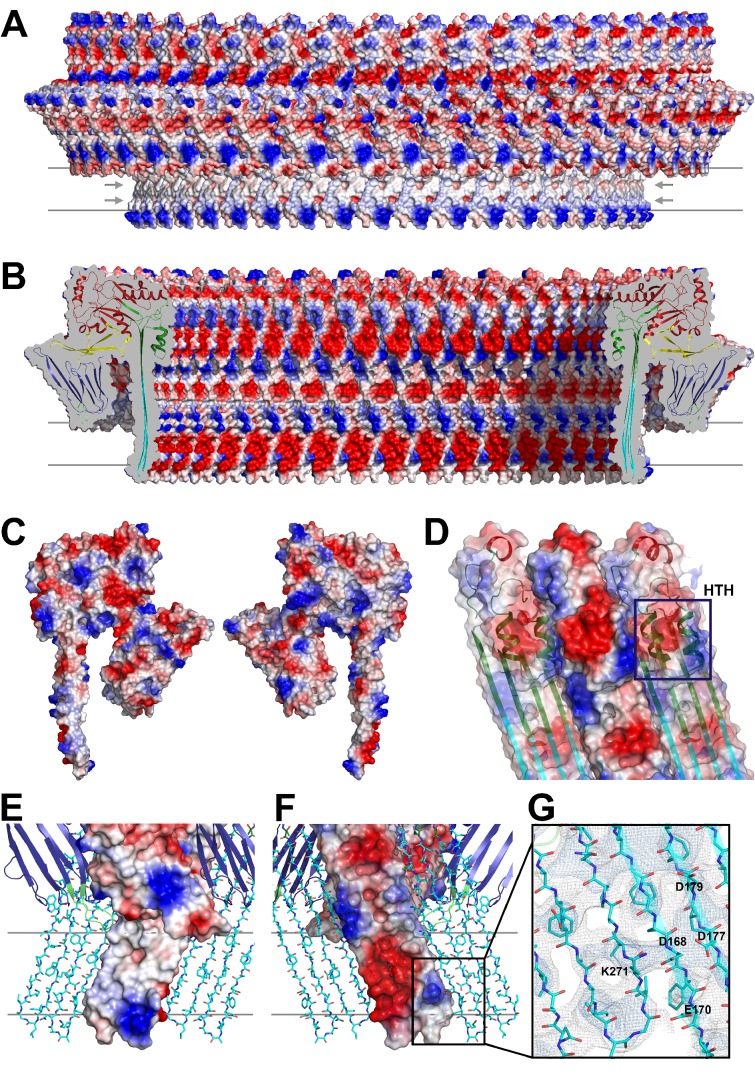
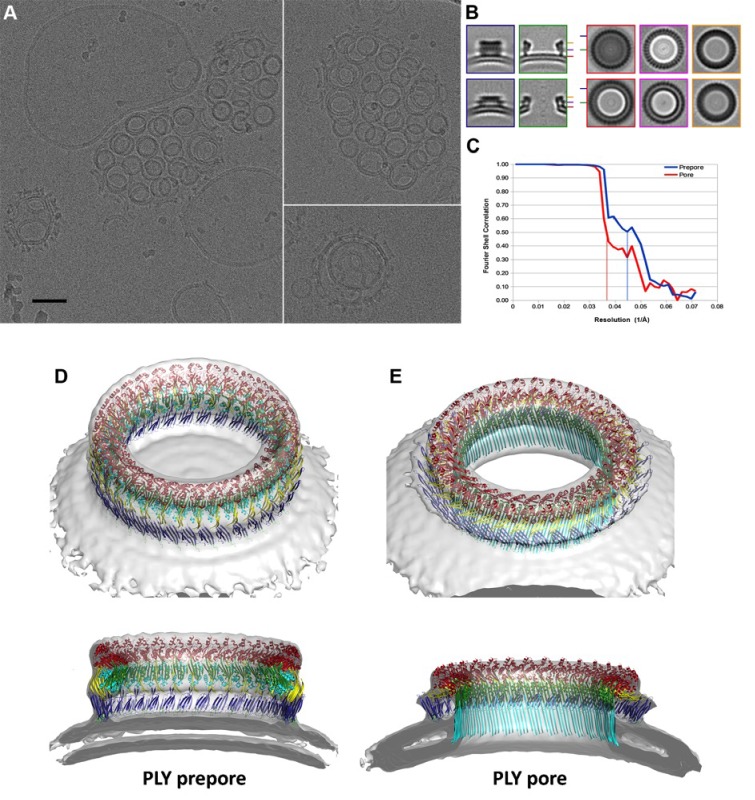
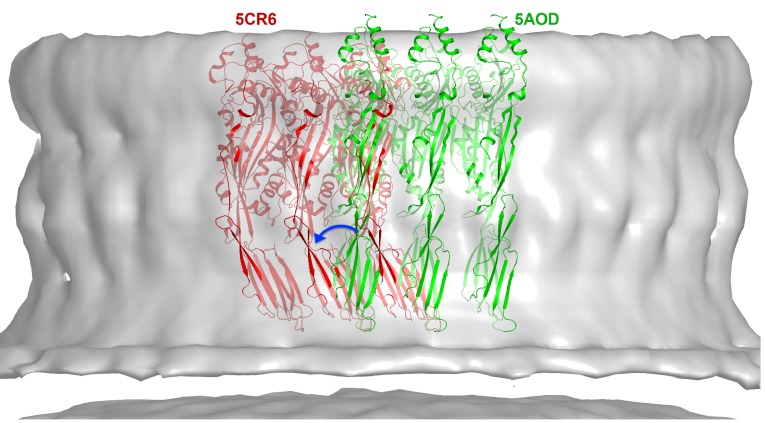
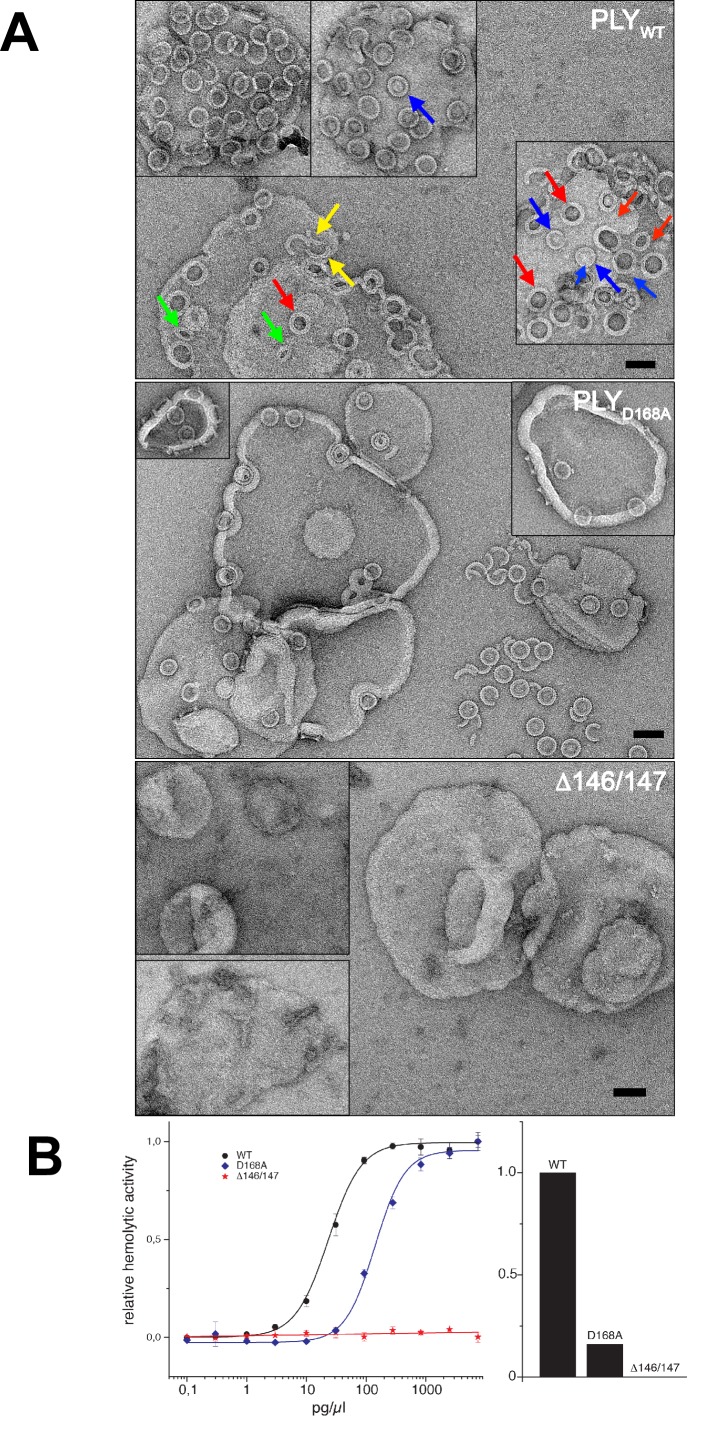
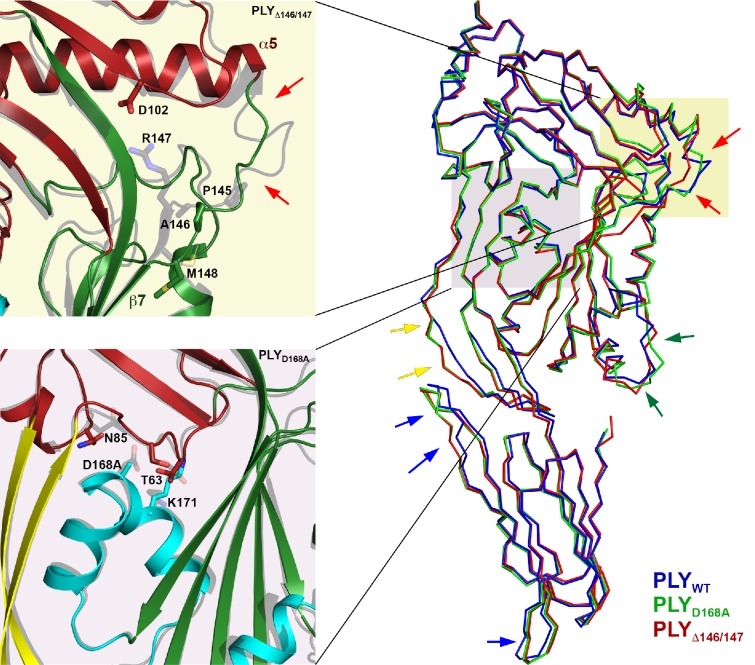
Many pathogenic bacteria produce pore-forming toxins to attack and kill human cells. We have determined the 4.5 Å structure of the ~2.2 MDa pore complex of pneumolysin, the main virulence factor of , by cryoEM. The pneumolysin pore is a 400 Å ring of 42 membrane-inserted monomers. Domain 3 of the soluble toxin refolds into two ~85 Å β-hairpins that traverse the lipid bilayer and assemble into a 168-strand β-barrel. The pore complex is stabilized by salt bridges between β-hairpins of adjacent subunits and an internal α-barrel. The apolar outer barrel surface with large sidechains is immersed in the lipid bilayer, while the inner barrel surface is highly charged. Comparison of the cryoEM pore complex to the prepore structure obtained by electron cryo-tomography and the x-ray structure of the soluble form reveals the detailed mechanisms by which the toxin monomers insert into the lipid bilayer to perforate the target membrane.
许多致病细菌会产生成孔毒素来攻击和杀死人体细胞。我们通过冷冻电镜确定了肺炎链球菌溶血素(肺炎链球菌的主要毒力因子)约2.2 MDa孔复合物的4.5 Å结构。肺炎链球菌溶血素孔是由42个插入膜中的单体组成的400 Å环。可溶性毒素的结构域3重折叠成两个约85 Å的β发夹结构,穿过脂质双层并组装成一个168链的β桶。孔复合物通过相邻亚基的β发夹结构与内部α桶之间的盐桥得以稳定。具有大侧链的非极性外桶表面浸入脂质双层中,而内桶表面则带有大量电荷。将冷冻电镜孔复合物与通过电子冷冻断层扫描获得的前孔结构以及可溶性形式的x射线结构进行比较,揭示了毒素单体插入脂质双层以穿透靶膜的详细机制。