Kohzuma Kaori, Froehlich John E, Davis Geoffry A, Temple Joshua A, Minhas Deepika, Dhingra Amit, Cruz Jeffrey A, Kramer David M
Department of Energy Plant Research Laboratory, Michigan State University, East LansingMI, United States.
Department of Biochemistry and Molecular Biology, Michigan State University, East LansingMI, United States.
Front Plant Sci. 2017 Jul 24;8:1248. doi: 10.3389/fpls.2017.01248. eCollection 2017.
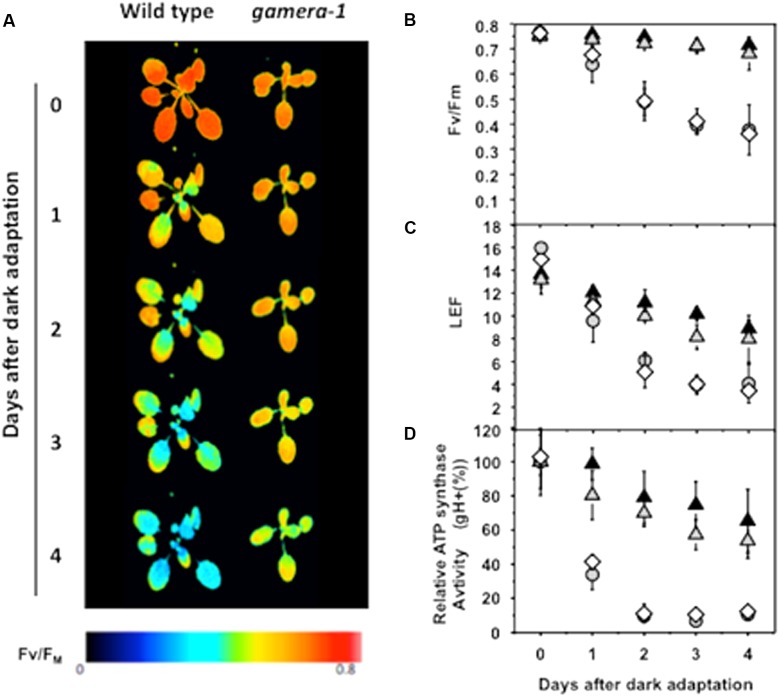
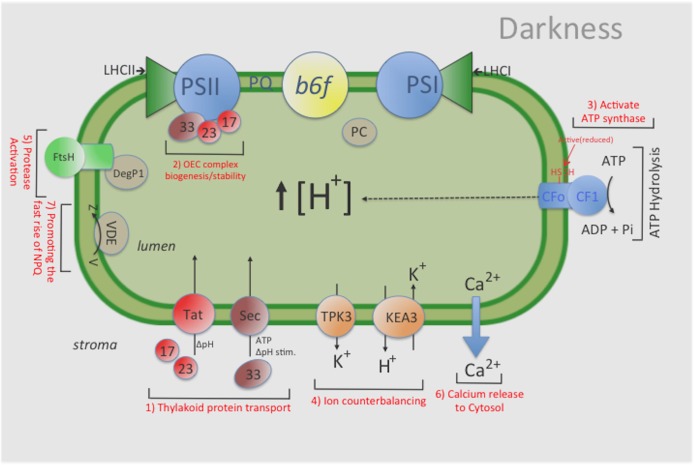
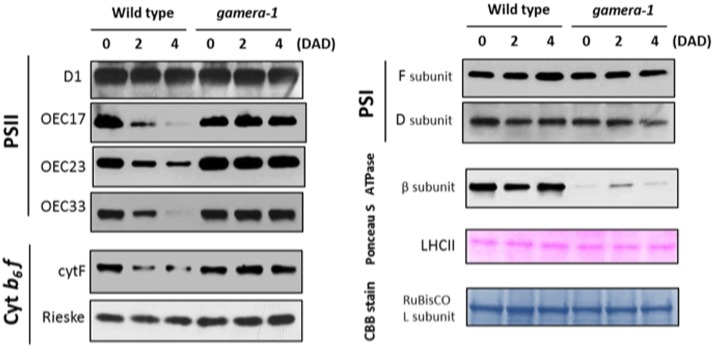
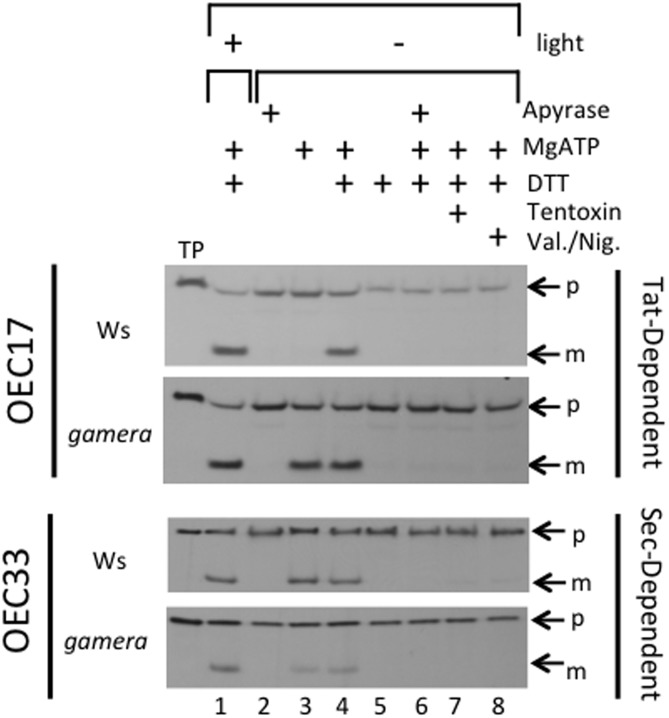
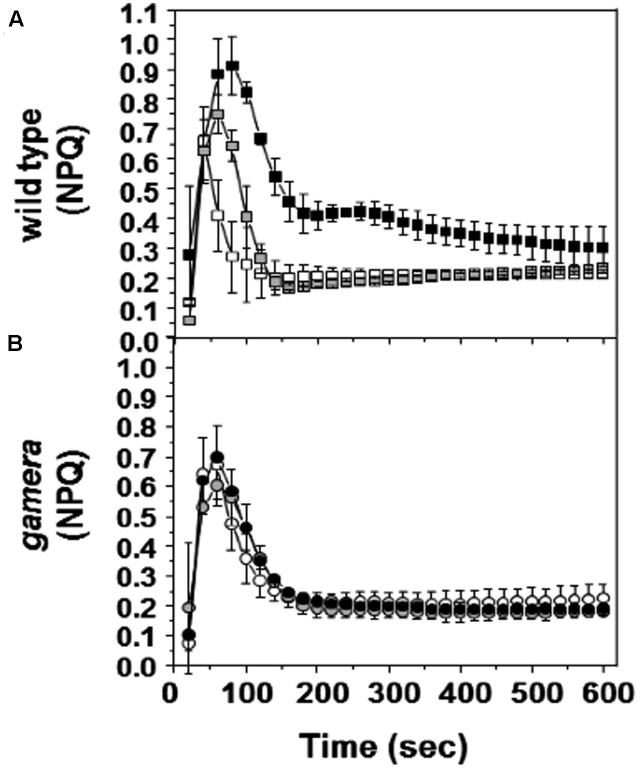
The chloroplast ATP synthase catalyzes the light-driven synthesis of ATP and is activated in the light and inactivated in the dark by redox-modulation through the thioredoxin system. It has been proposed that this down-regulation is important for preventing wasteful hydrolysis of ATP in the dark. To test this proposal, we compared the effects of extended dark exposure in Arabidopsis lines expressing the wild-type and mutant forms of ATP synthase that are redox regulated or constitutively active. In contrast to the predictions of the model, we observed that plants with wild-type redox regulation lost photosynthetic capacity rapidly in darkness, whereas those expressing redox-insensitive form were far more stable. To explain these results, we propose that in wild-type plants, down-regulation of ATP synthase inhibits ATP hydrolysis, leading to dissipation of thylakoid proton motive force (pmf) and subsequent inhibition of protein transport across the thylakoid through the twin arginine transporter (Tat)-dependent and Sec-dependent import pathways, resulting in the selective loss of specific protein complexes. By contrast, in mutants with a redox-insensitive ATP synthase, pmf is maintained by ATP hydrolysis, thus allowing protein transport to maintain photosynthetic activities for extended periods in the dark. Hence, a basal level of Tat-dependent, as well as, Sec-dependent import activity, in the dark helps replenishes certain components of the photosynthetic complexes and thereby aids in maintaining overall complex activity. However, the influence of a dark pmf on thylakoid protein import, by itself, could not explain all the effects we observed in this study. For example, we also observed in wild type plants a large transient buildup of thylakoid pmf and nonphotochemical exciton quenching upon sudden illumination of dark adapted plants. Therefore, we conclude that down-regulation of the ATP synthase is probably not related to preventing loss of ATP . Instead, ATP synthase redox regulation may be impacting a number of cellular processes such as (1) the accumulation of chloroplast proteins and/or ions or (2) the responses of photosynthesis to rapid changes in light intensity. A model highlighting the complex interplay between ATP synthase regulation and pmf in maintaining various chloroplast functions in the dark is presented. We uncover an unexpected role for thioredoxin modulation of the chloroplast ATP synthase in regulating the dark-stability of the photosynthetic apparatus, most likely by controlling thylakoid membrane transport of proteins and ions.
叶绿体ATP合酶催化由光驱动的ATP合成,通过硫氧还蛋白系统进行氧化还原调节,在光照下被激活,在黑暗中失活。有人提出,这种下调对于防止黑暗中ATP的浪费性水解很重要。为了验证这一观点,我们比较了在表达野生型和突变型ATP合酶的拟南芥品系中延长黑暗暴露的影响,这些ATP合酶是氧化还原调节的或组成型活性的。与模型预测相反,我们观察到具有野生型氧化还原调节的植物在黑暗中迅速丧失光合能力,而那些表达对氧化还原不敏感形式的植物则稳定得多。为了解释这些结果,我们提出,在野生型植物中,ATP合酶的下调抑制ATP水解,导致类囊体质子动力势(pmf)的耗散,随后抑制蛋白质通过双精氨酸转运蛋白(Tat)依赖性和Sec依赖性导入途径跨类囊体的转运,导致特定蛋白质复合物的选择性丧失。相比之下,在具有对氧化还原不敏感的ATP合酶的突变体中,pmf通过ATP水解得以维持,从而使蛋白质转运在黑暗中长时间维持光合活性。因此,黑暗中基础水平的Tat依赖性以及Sec依赖性导入活性有助于补充光合复合物的某些成分,从而有助于维持整体复合物活性。然而,黑暗中的pmf对类囊体蛋白质导入本身的影响并不能解释我们在本研究中观察到的所有效应。例如,我们还在野生型植物中观察到,暗适应植物突然光照后,类囊体pmf和非光化学激子猝灭会出现大量瞬时积累。因此,我们得出结论,ATP合酶的下调可能与防止ATP损失无关。相反,ATP合酶的氧化还原调节可能会影响许多细胞过程,例如(1)叶绿体蛋白质和/或离子的积累,或(2)光合作用对光强快速变化的响应。本文提出了一个模型,突出了ATP合酶调节与pmf在黑暗中维持各种叶绿体功能之间的复杂相互作用。我们揭示了硫氧还蛋白对叶绿体ATP合酶的调节在调节光合装置的黑暗稳定性方面的一个意想不到的作用,很可能是通过控制类囊体膜上蛋白质和离子的转运来实现的。