Biophysics Graduate Group, University of California, Berkeley, Berkeley, United States.
Physics Department, University of California, Berkeley, Berkeley, United States.
Elife. 2017 Sep 20;6:e28606. doi: 10.7554/eLife.28606.
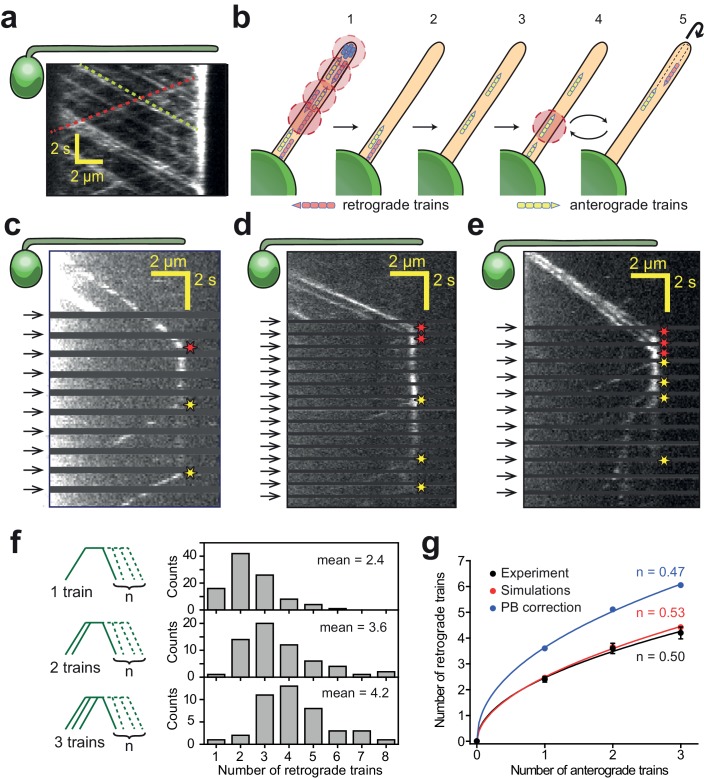
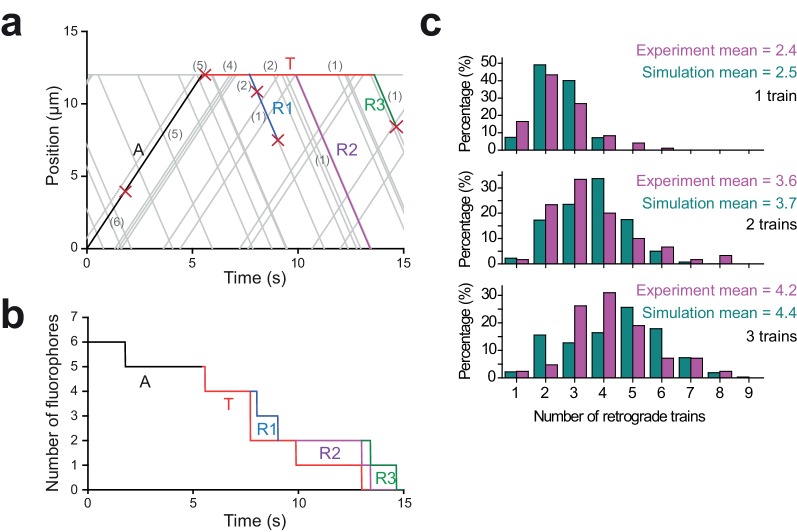
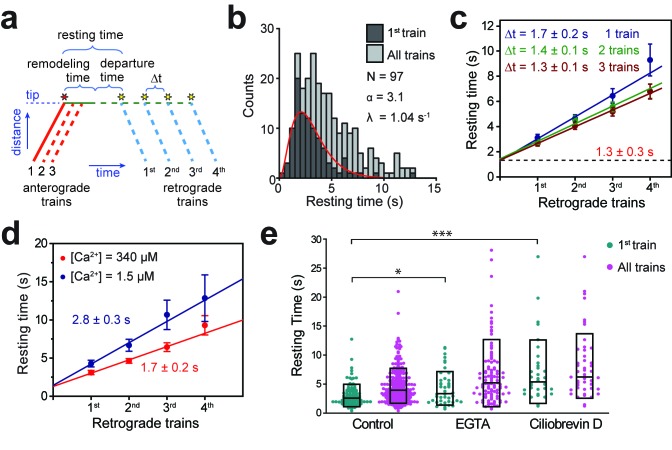
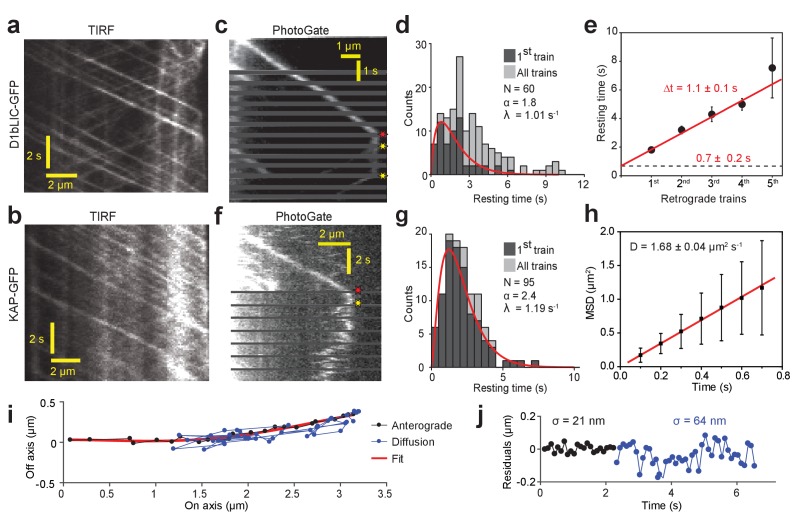
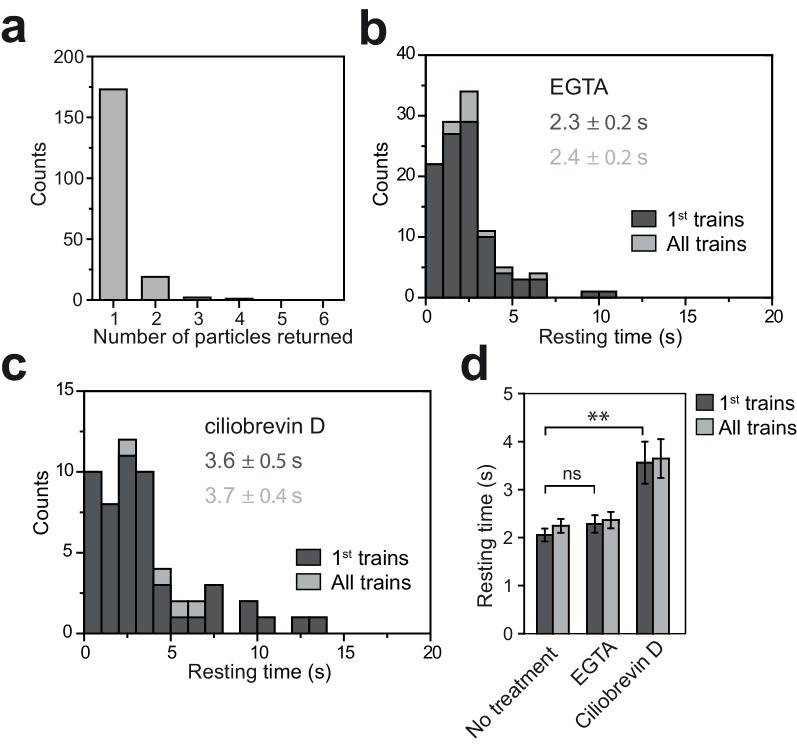
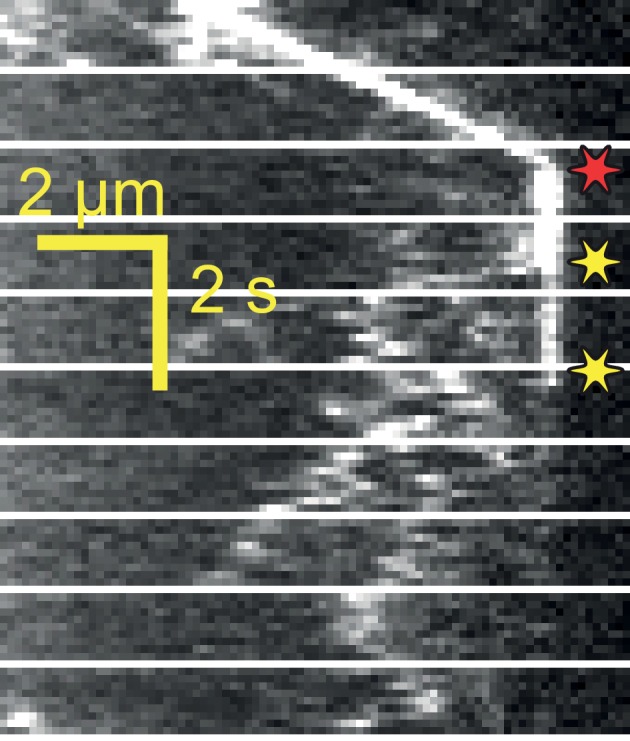
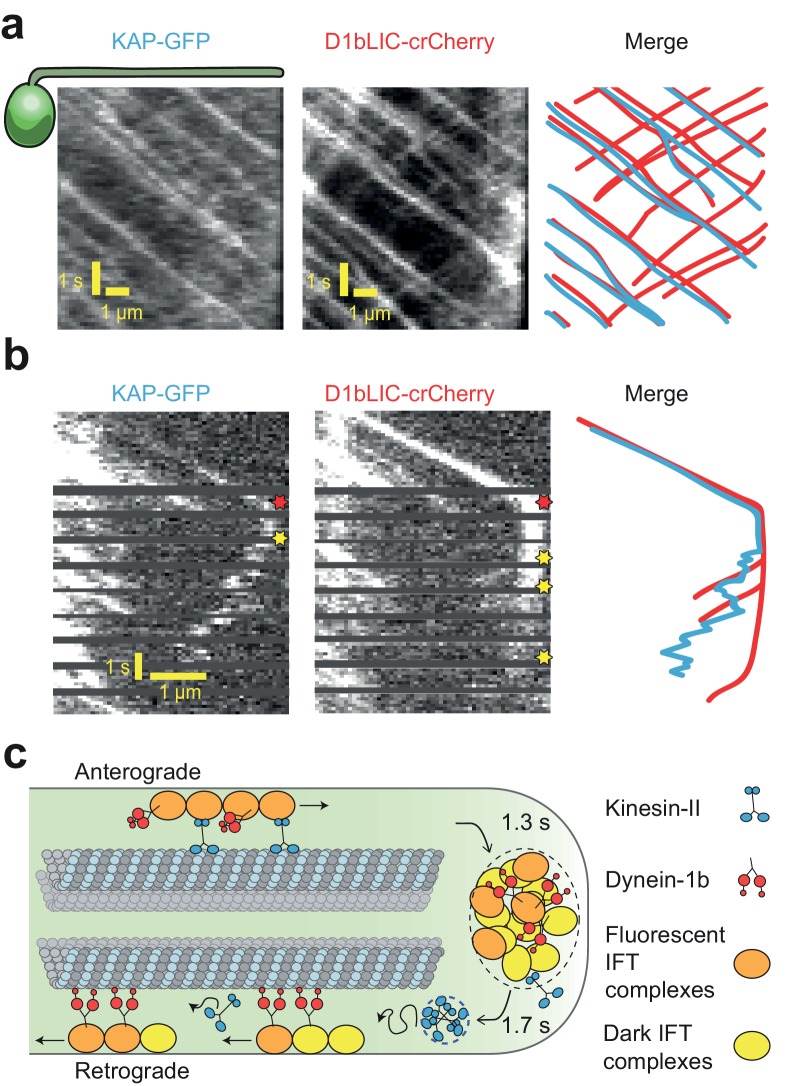
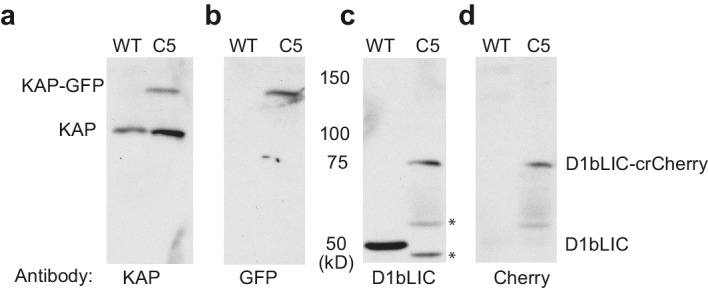
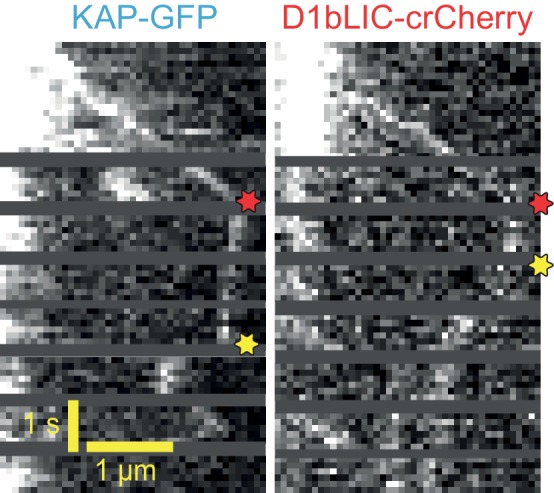
Intraflagellar transport (IFT) is essential for the elongation and maintenance of eukaryotic cilia and flagella. Due to the traffic jam of multiple trains at the ciliary tip, how IFT trains are remodeled in these turnaround zones cannot be determined by conventional imaging. Using PhotoGate, we visualized the full range of movement of single IFT trains and motors in flagella. Anterograde trains split apart and IFT complexes mix with each other at the tip to assemble retrograde trains. Dynein-1b is carried to the tip by kinesin-II as inactive cargo on anterograde trains. Unlike dynein-1b, kinesin-II detaches from IFT trains at the tip and diffuses in flagella. As the flagellum grows longer, diffusion delays return of kinesin-II to the basal body, depleting kinesin-II available for anterograde transport. Our results suggest that dissociation of kinesin-II from IFT trains serves as a negative feedback mechanism that facilitates flagellar length control in .
纤毛和鞭毛的伸长和维持需要内鞭毛运输(IFT)。由于在纤毛尖端有多列火车的交通堵塞,因此无法通过常规成像确定 IFT 列车在这些周转区如何进行重塑。使用 PhotoGate,我们可视化了鞭毛中单列 IFT 列车和马达的全范围运动。向前行驶的列车分开,IFT 复合物在尖端相互混合以组装逆行列车。动力蛋白-1b 作为向前行驶列车上的无活性货物被 kinesin-II 携带到尖端。与动力蛋白-1b 不同,kinesin-II 在尖端从 IFT 列车上脱离并在鞭毛中扩散。随着鞭毛变长,扩散延迟使 kinesin-II 回到基体,耗尽了用于向前运输的 kinesin-II。我们的结果表明,kinesin-II 从 IFT 列车上解离充当了负反馈机制,有助于在 中控制鞭毛长度。