Department of Biology, West Virginia University, Morgantown, West Virginia, USA.
Department of Microbiology and Immunology, University of Maryland School of Medicine, Baltimore, Maryland, USA.
mBio. 2017 Sep 26;8(5):e00859-17. doi: 10.1128/mBio.00859-17.
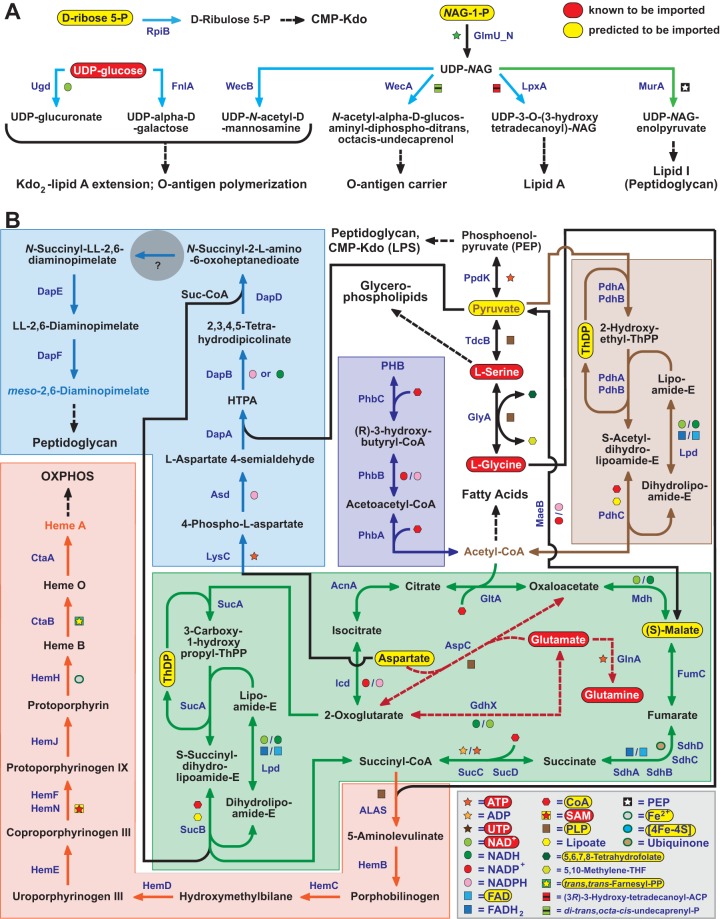
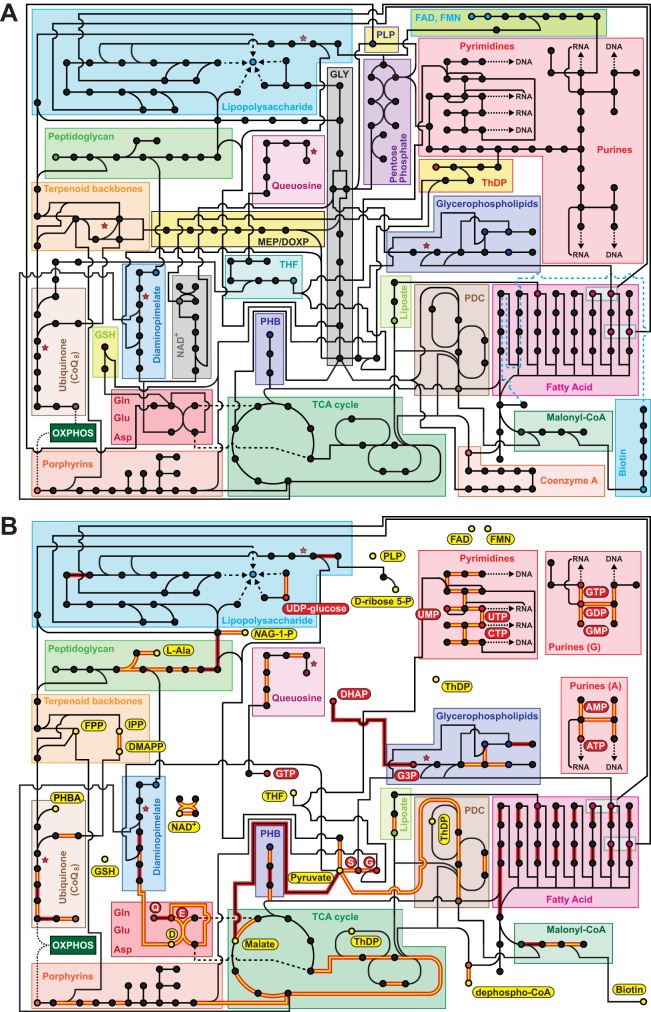
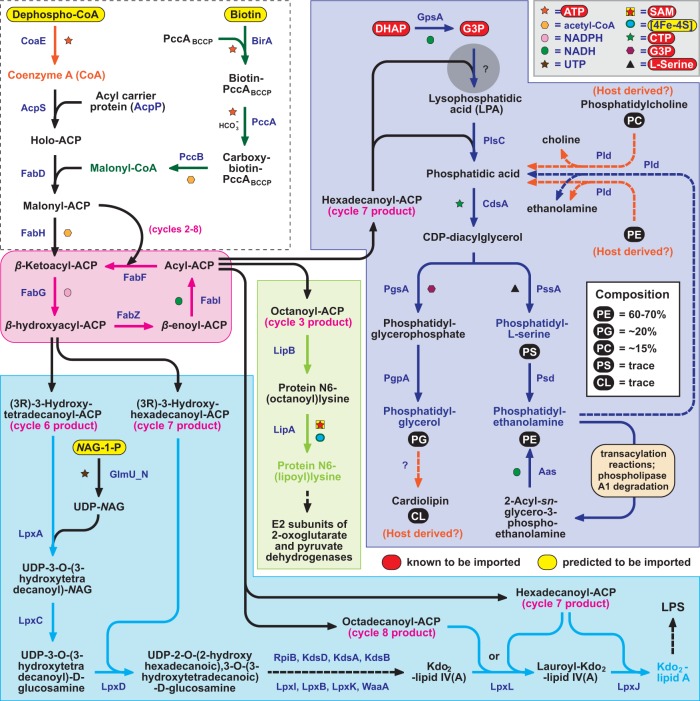
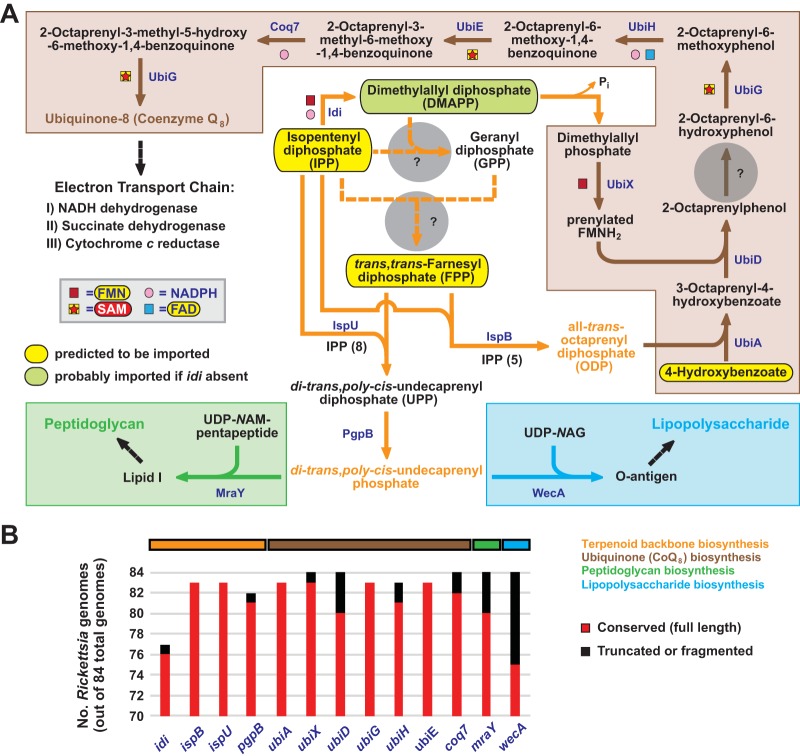
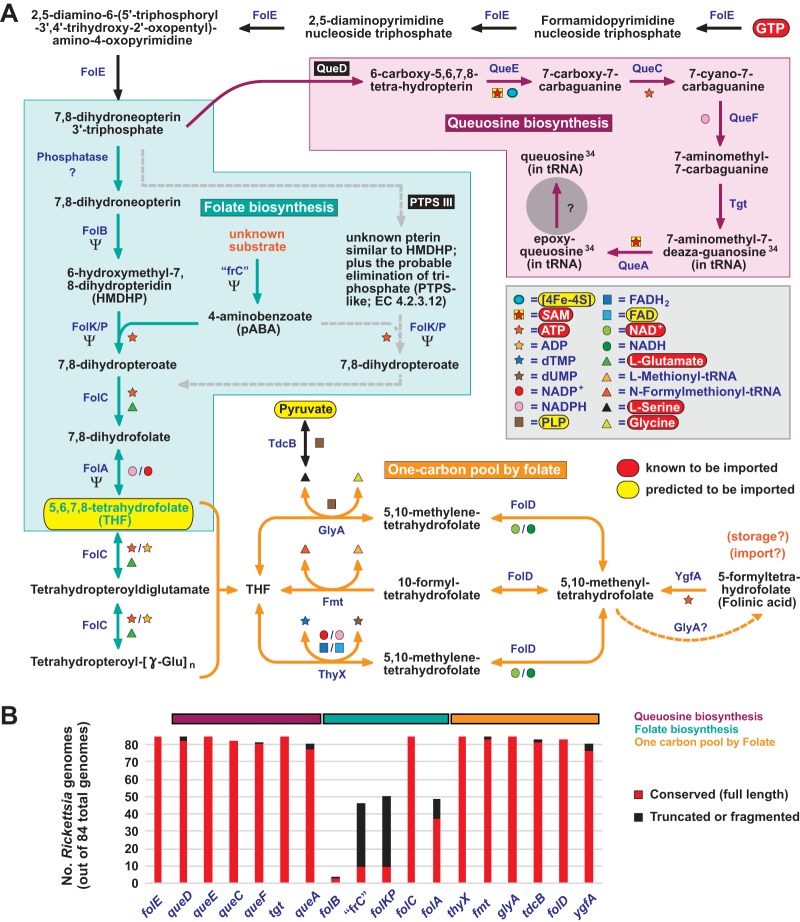
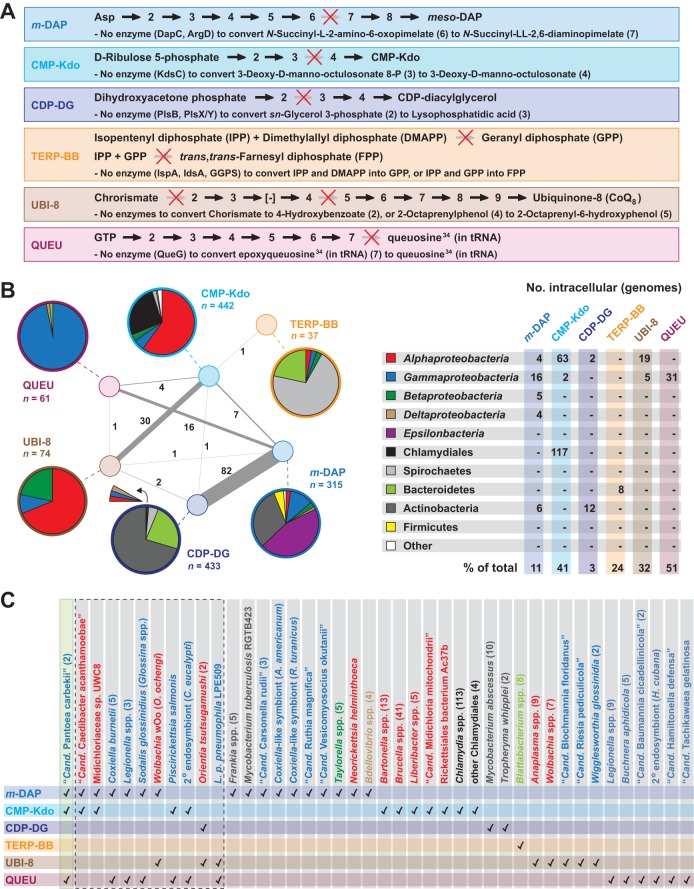
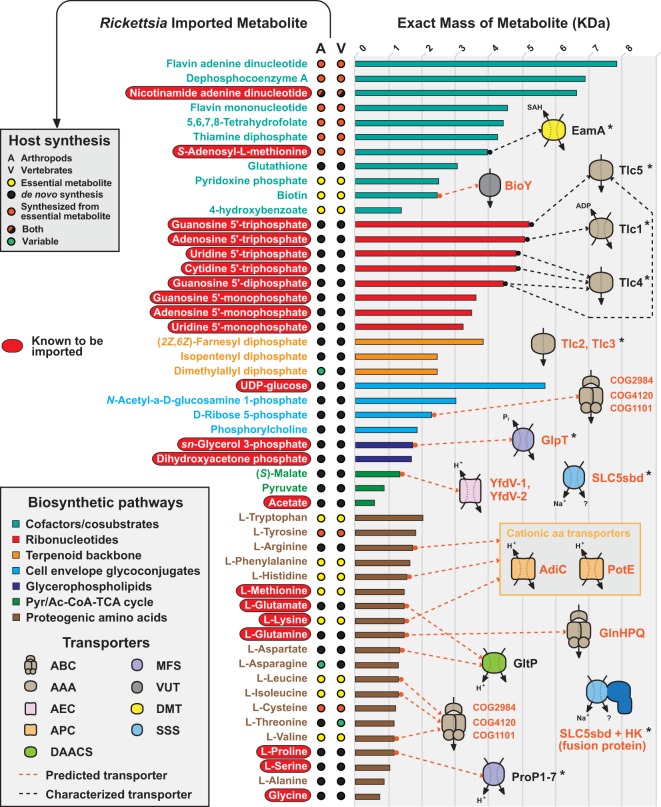
Reductive genome evolution has purged many metabolic pathways from obligate intracellular (; ). While some aspects of host-dependent rickettsial metabolism have been characterized, the array of host-acquired metabolites and their cognate transporters remains unknown. This dearth of information has thwarted efforts to obtain an axenic culture, a major impediment to conventional genetic approaches. Using phylogenomics and computational pathway analysis, we reconstructed the metabolic and transport network, identifying 51 host-acquired metabolites (only 21 previously characterized) needed to compensate for degraded biosynthesis pathways. In the absence of glycolysis and the pentose phosphate pathway, cell envelope glycoconjugates are synthesized from three imported host sugars, with a range of additional host-acquired metabolites fueling the tricarboxylic acid cycle. Fatty acid and glycerophospholipid pathways also initiate from host precursors, and import of both isoprenes and terpenoids is required for the synthesis of ubiquinone and the lipid carrier of lipid I and O-antigen. Unlike metabolite-provisioning bacterial symbionts of arthropods, rickettsiae cannot synthesize B vitamins or most other cofactors, accentuating their parasitic nature. Six biosynthesis pathways contain holes (missing enzymes); similar patterns in taxonomically diverse bacteria suggest alternative enzymes that await discovery. A paucity of characterized and predicted transporters emphasizes the knowledge gap concerning how rickettsiae import host metabolites, some of which are large and not known to be transported by bacteria. Collectively, our reconstructed metabolic network offers clues to how rickettsiae hijack host metabolic pathways. This blueprint for growth determinants is an important step toward the design of axenic media to rescue rickettsiae from the eukaryotic cell. A hallmark of obligate intracellular bacteria is the tradeoff of metabolic genes for the ability to acquire host metabolites. For species of , arthropod-borne parasites with the potential to cause serious human disease, the range of pilfered host metabolites is unknown. This information is critical for dissociating rickettsiae from eukaryotic cells to facilitate rickettsial genetic manipulation. In this study, we reconstructed the metabolic network and identified 51 host metabolites required to compensate patchwork biosynthesis pathways. Remarkably, some metabolites are not known to be transported by any bacteria, and overall, few cognate transporters were identified. Several pathways contain missing enzymes, yet similar pathways in unrelated bacteria indicate convergence and possible novel enzymes awaiting characterization. Our work illuminates the parasitic nature by which rickettsiae hijack host metabolism to counterbalance numerous disintegrated biosynthesis pathways that have arisen through evolution within the eukaryotic cell. This metabolic blueprint reveals what a axenic medium might entail.
专性细胞内寄生菌()已经通过还原基因组进化消除了许多代谢途径。虽然宿主依赖的立克次体代谢的某些方面已经得到了描述,但宿主获得的代谢物及其相应的转运蛋白的范围仍然未知。这种信息的缺乏阻碍了获得无菌培养物的努力,这是传统遗传方法的主要障碍。我们使用系统发育基因组学和计算途径分析,重建了代谢和运输网络,确定了 51 种需要补偿降解生物合成途径的宿主获得代谢物(仅以前鉴定了 21 种)。在没有糖酵解和戊糖磷酸途径的情况下,细胞包膜糖缀合物是由三种进口的宿主糖合成的,还有一系列其他的宿主获得代谢物为三羧酸循环提供燃料。脂肪酸和甘油磷脂途径也从宿主前体开始,并且需要导入异戊烯和萜烯来合成泛醌和脂质 I 和 O-抗原的脂质载体。与节肢动物的提供代谢物的细菌共生体不同,立克次氏体不能合成 B 族维生素或大多数其他辅因子,这突出了它们的寄生性质。六个生物合成途径存在缺口(缺少酶);在分类上多样化的细菌中存在类似的模式表明有待发现的替代酶。特征描述和预测的转运蛋白数量较少强调了有关立克次氏体如何导入宿主代谢物的知识空白,其中一些代谢物较大且未知是否被细菌转运。总的来说,我们重建的代谢网络提供了线索,说明立克次氏体如何劫持宿主代谢途径。这个生长决定因素的蓝图是设计无菌培养基以将立克次氏体从真核细胞中解救出来的重要一步。专性细胞内细菌的一个标志是代谢基因与获取宿主代谢物的能力之间的权衡。对于节肢动物传播的寄生虫种,具有引起严重人类疾病的潜力,被盗用的宿主代谢物范围尚不清楚。这些信息对于将立克次氏体与真核细胞分离以促进立克次氏体遗传操作至关重要。在这项研究中,我们重建了代谢网络,并确定了 51 种宿主代谢物,这些代谢物需要补偿拼凑的生物合成途径。值得注意的是,一些代谢物已知不是由任何细菌转运的,而且总的来说,鉴定出的同源转运蛋白很少。一些途径缺少酶,但在不相关的细菌中类似的途径表明趋同,可能有新的酶有待鉴定。我们的工作阐明了立克次氏体通过劫持宿主代谢来平衡在真核细胞内进化过程中出现的许多瓦解的生物合成途径的寄生性质。这个代谢蓝图揭示了一个无菌培养基可能需要什么。