Institute of Pharmacology and Toxicology, University of Zurich, Winterthurerstrasse 190, 8057, Zurich, Switzerland.
Laboratory for Animal Model Pathology (LAMP), Institute of Veterinary Pathology, University of Zurich, Winterthurerstrasse 268, 8057, Zurich, Switzerland.
Arch Toxicol. 2017 Nov;91(11):3571-3585. doi: 10.1007/s00204-017-2060-4. Epub 2017 Oct 3.
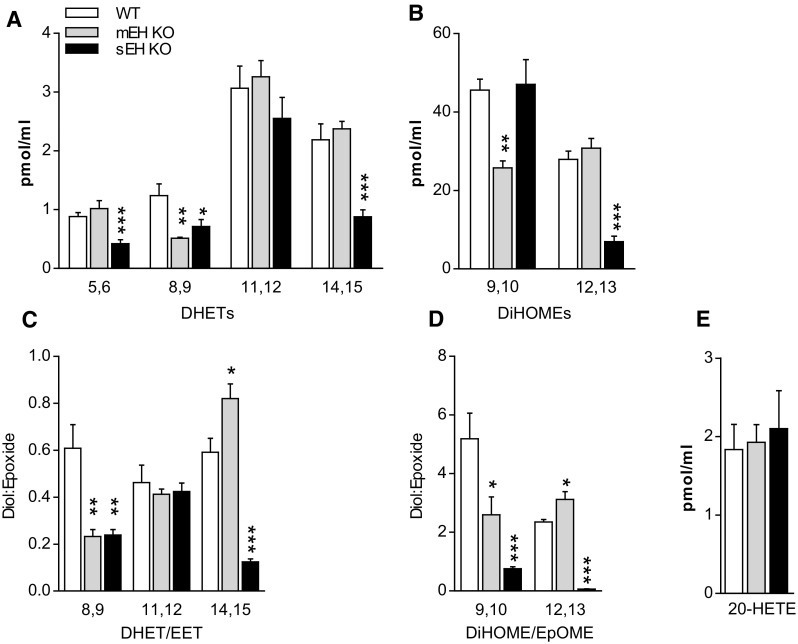
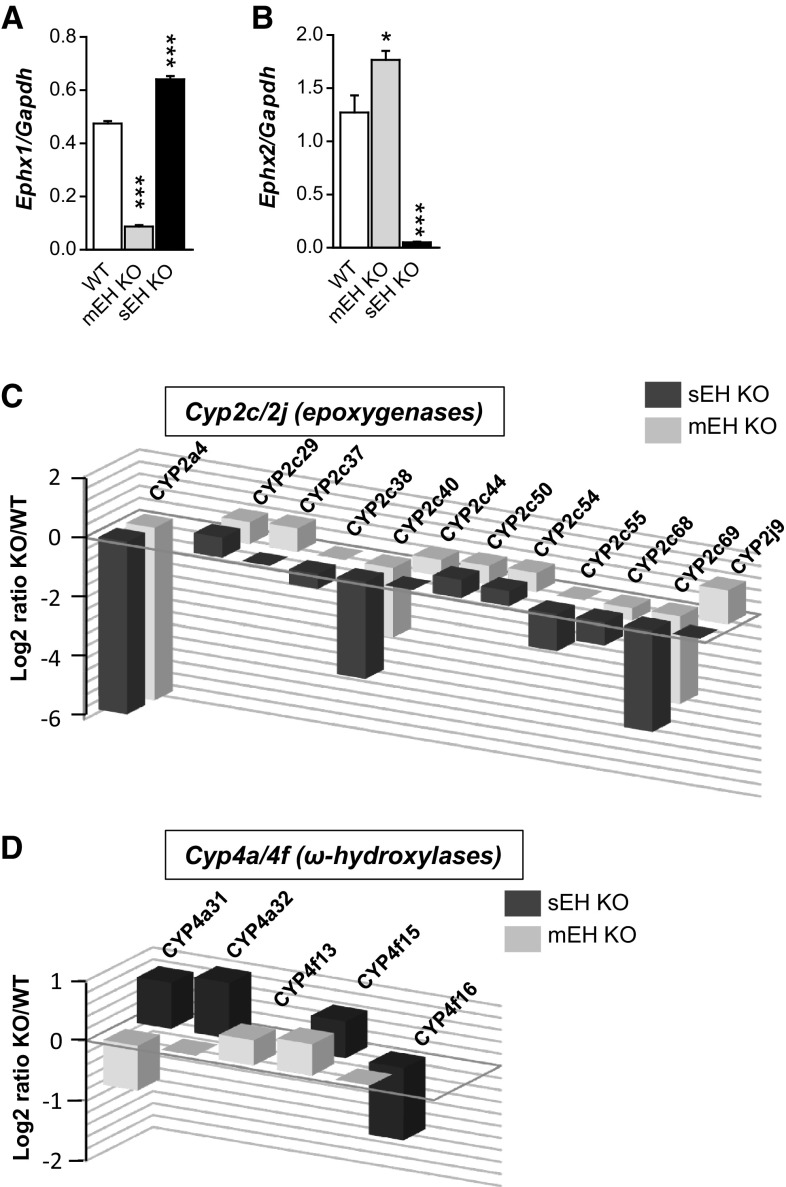
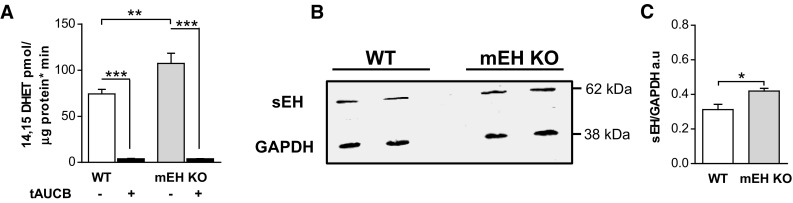
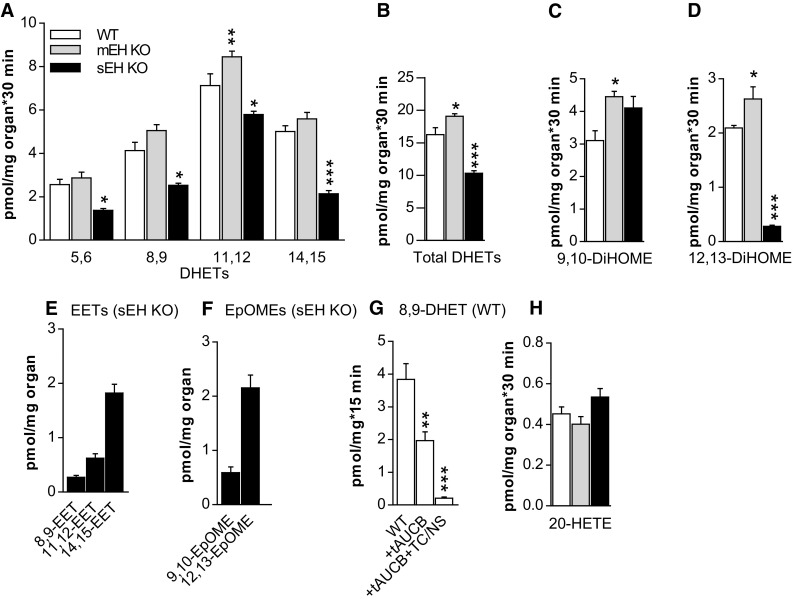
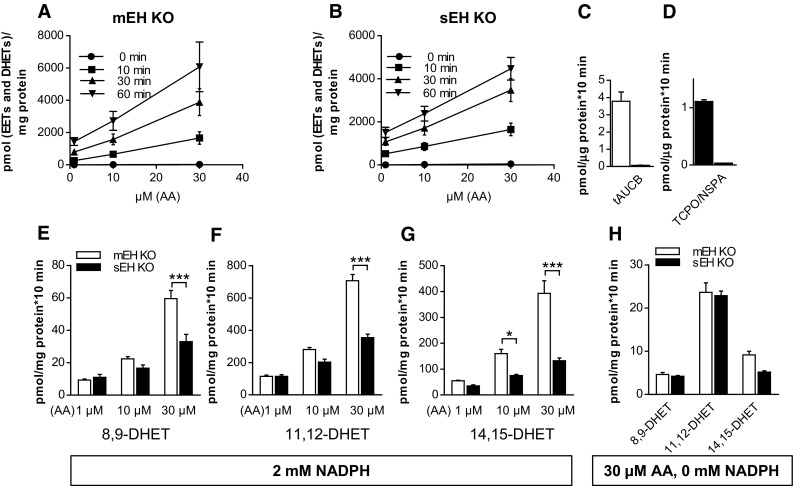
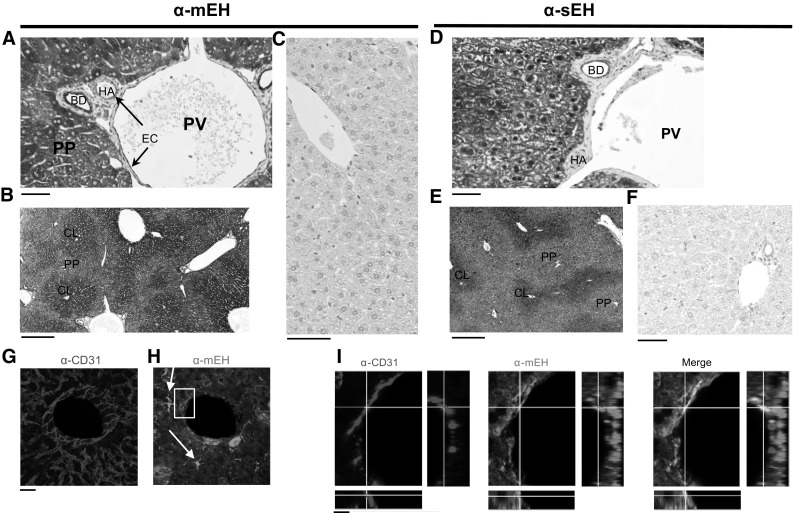
Microsomal and soluble epoxide hydrolase (mEH and sEH) fulfill apparently distinct roles: Whereas mEH detoxifies xenobiotics, sEH hydrolyzes fatty acid (FA) signaling molecules and is thus implicated in a variety of physiological functions. These epoxy FAs comprise epoxyeicosatrienoic acids (EETs) and epoxy-octadecenoic acids (EpOMEs), which are formed by CYP epoxygenases from arachidonic acid (AA) and linoleic acid, respectively, and then are hydrolyzed to their respective diols, the so-called DHETs and DiHOMEs. Although EETs and EpOMEs are also substrates for mEH, its role in lipid signaling is considered minor due to lower abundance and activity relative to sEH. Surprisingly, we found that in plasma from mEH KO mice, hydrolysis rates for 8,9-EET and 9,10-EpOME were reduced by 50% compared to WT plasma. This strongly suggests that mEH contributes substantially to the turnover of these FA epoxides-despite kinetic parameters being in favor of sEH. Given the crucial role of liver in controlling plasma diol levels, we next studied the capacity of sEH and mEH KO liver microsomes to synthesize DHETs with varying concentrations of AA (1-30 μM) and NADPH. mEH-generated DHET levels were similar to the ones generated by sEH, when AA concentrations were low (1 μM) or epoxygenase activity was curbed by modulating NADPH. With increasing AA concentrations sEH became more dominant and with 30 μM AA produced twice the level of DHETs compared to mEH. Immunohistochemistry of C57BL/6 liver slices further revealed that mEH expression was more widespread than sEH expression. mEH immunoreactivity was detected in hepatocytes, Kupffer cells, endothelial cells, and bile duct epithelial cells, while sEH immunoreactivity was confined to hepatocytes and bile duct epithelial cells. Finally, transcriptome analysis of WT, mEH KO, and sEH KO liver was carried out to discern transcriptional changes associated with the loss of EH genes along the CYP-epoxygenase-EH axis. We found several prominent dysregulations occurring in a parallel manner in both KO livers: (a) gene expression of Ephx1 (encoding for mEH protein) was increased 1.35-fold in sEH KO, while expression of Ephx2 (encoding for sEH protein) was increased 1.4-fold in mEH KO liver; (b) Cyp2c genes, encoding for the predominant epoxygenases in mouse liver, were mostly dysregulated in the same manner in both sEH and mEH KO mice, showing that loss of either EH has a similar impact. Taken together, mEH appears to play a leading role in the hydrolysis of 8,9-EET and 9,10-EpOME and also contributes to the hydrolysis of other FA epoxides. It probably profits from its high affinity for FA epoxides under non-saturating conditions and its close physical proximity to CYP epoxygenases, and compensates its lower abundance by a more widespread expression, being the only EH present in several sEH-lacking cell types.
微粒体和可溶性环氧化物水解酶(mEH 和 sEH)发挥着明显不同的作用:mEH 解毒外来化合物,而 sEH 水解脂肪酸(FA)信号分子,因此参与多种生理功能。这些环氧 FA 包括环氧二十碳三烯酸(EETs)和环氧十八碳烯酸(EpOMEs),它们分别由细胞色素 P450 环氧化物酶从花生四烯酸(AA)和亚油酸中形成,然后水解为各自的二醇,即所谓的 DHETs 和 DiHOMEs。尽管 EETs 和 EpOMEs 也是 mEH 的底物,但由于其丰度和活性相对于 sEH 较低,其在脂质信号中的作用被认为较小。令人惊讶的是,我们发现 mEH KO 小鼠的血浆中,8,9-EET 和 9,10-EpOME 的水解率比 WT 血浆降低了 50%。这强烈表明 mEH 对这些 FA 环氧的周转做出了实质性贡献-尽管动力学参数有利于 sEH。鉴于肝脏在控制血浆二醇水平方面的关键作用,我们接下来研究了 sEH 和 mEH KO 肝微粒体在不同 AA(1-30 μM)和 NADPH 浓度下合成 DHET 的能力。当 AA 浓度较低(1 μM)或通过调节 NADPH 抑制环氧酶活性时,mEH 产生的 DHET 水平与 sEH 产生的水平相似。随着 AA 浓度的增加,sEH 变得更加占主导地位,并且 30 μM AA 产生的 DHET 水平是 mEH 的两倍。C57BL/6 肝切片的免疫组织化学进一步表明,mEH 的表达比 sEH 的表达更为广泛。mEH 免疫反应性在肝细胞、库普弗细胞、内皮细胞和胆管上皮细胞中均有检测到,而 sEH 免疫反应性仅局限于肝细胞和胆管上皮细胞。最后,对 WT、mEH KO 和 sEH KO 肝脏进行了转录组分析,以辨别与 CYP-环氧酶-EH 轴上 EH 基因缺失相关的转录变化。我们发现两种 KO 肝脏中以平行方式发生了几种明显的失调:(a)sEH KO 中 Ephx1(编码 mEH 蛋白)的基因表达增加了 1.35 倍,而 mEH KO 肝脏中 Ephx2(编码 sEH 蛋白)的基因表达增加了 1.4 倍;(b)编码小鼠肝脏中主要环氧酶的 Cyp2c 基因在两种 sEH 和 mEH KO 小鼠中以相似的方式失调,表明任何一种 EH 的缺失都有类似的影响。总之,mEH 似乎在 8,9-EET 和 9,10-EpOME 的水解中发挥主导作用,并且还有助于其他 FA 环氧的水解。它可能受益于其在非饱和条件下对 FA 环氧的高亲和力及其与 CYP 环氧化物酶的紧密物理接近性,并通过更广泛的表达来弥补其丰度较低的不足,是几种缺乏 sEH 的细胞类型中唯一存在的 EH。