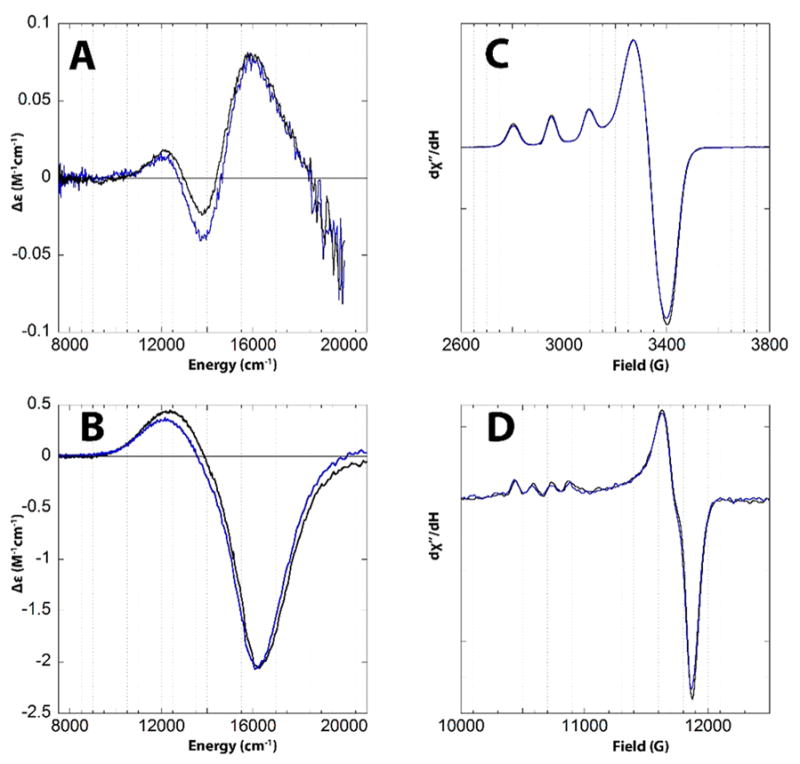
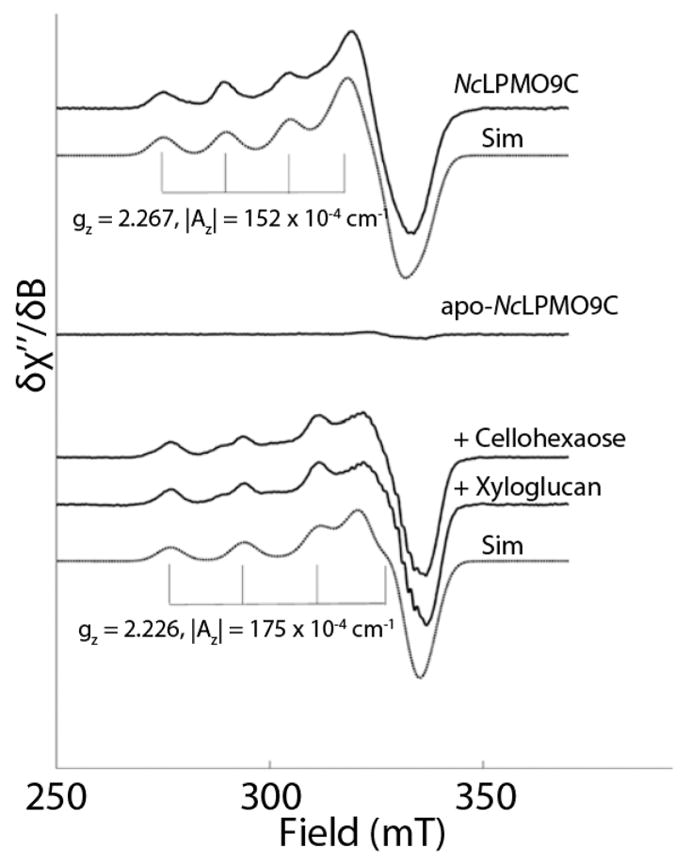
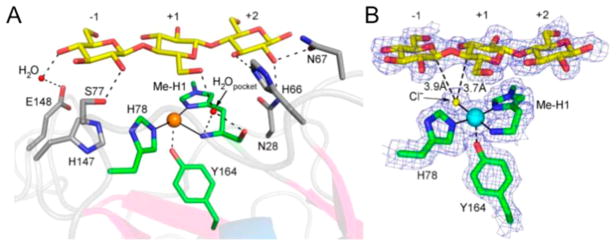
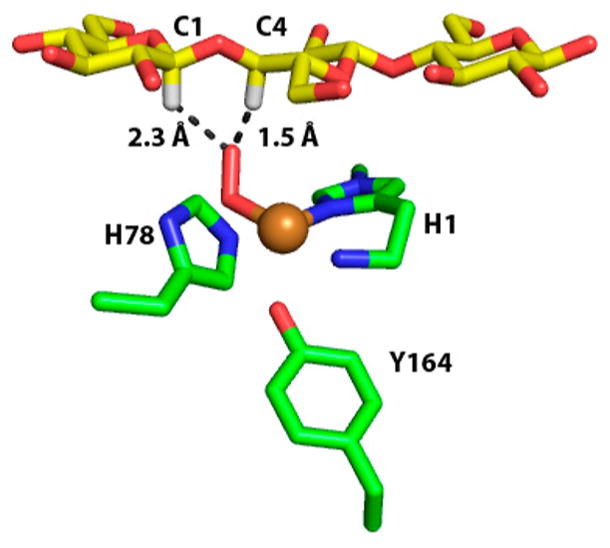
Department of Chemistry , Stanford University , Stanford , California 94305 , United States.
DuPont Industrial Biosciences , 925 Page Mill Road , Palo Alto , California 94304 , United States.
Chem Rev. 2018 Mar 14;118(5):2593-2635. doi: 10.1021/acs.chemrev.7b00421. Epub 2017 Nov 20.
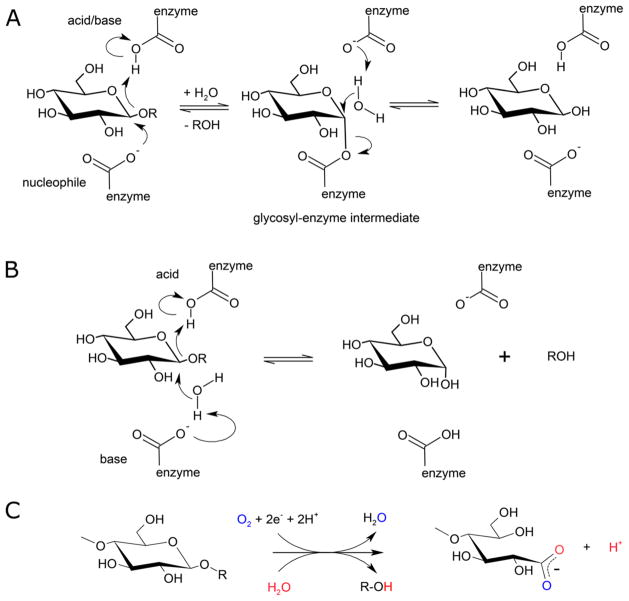
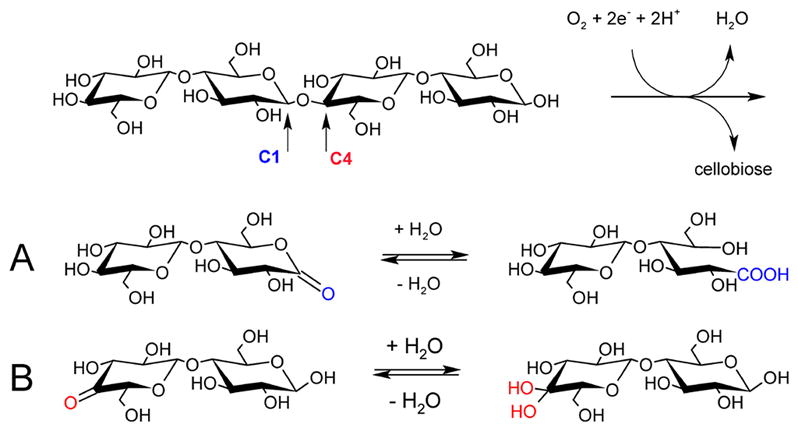
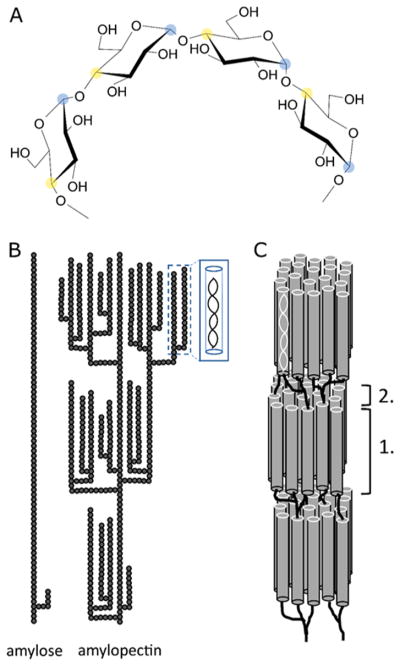
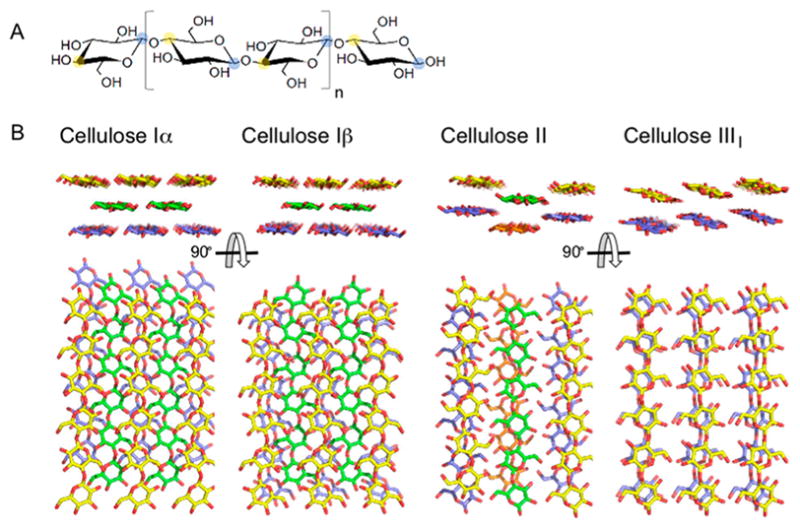
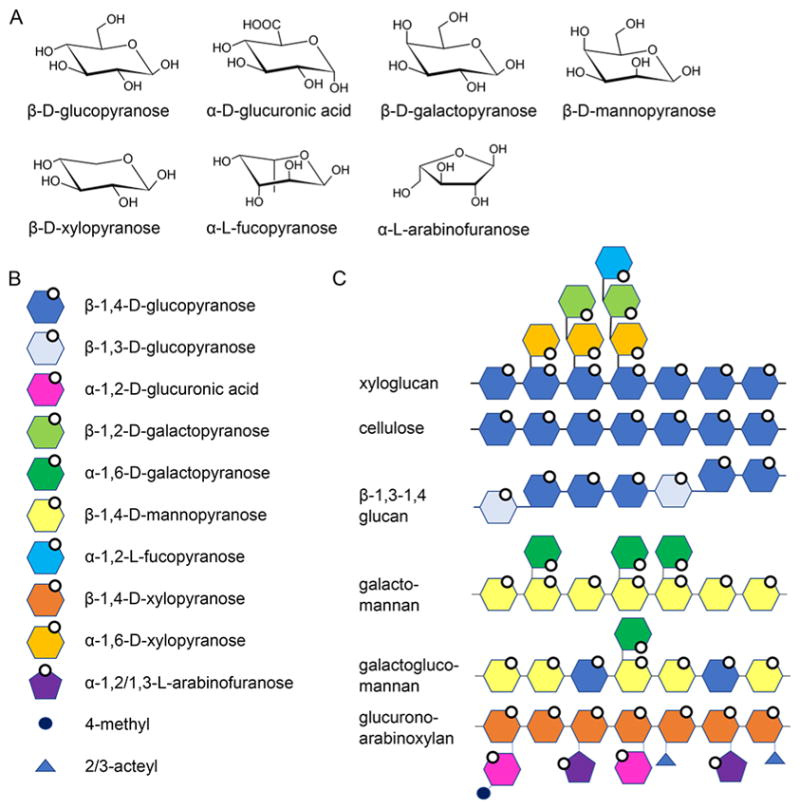
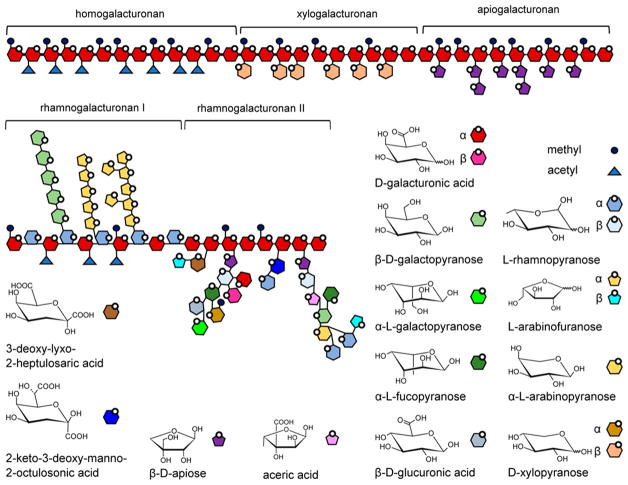
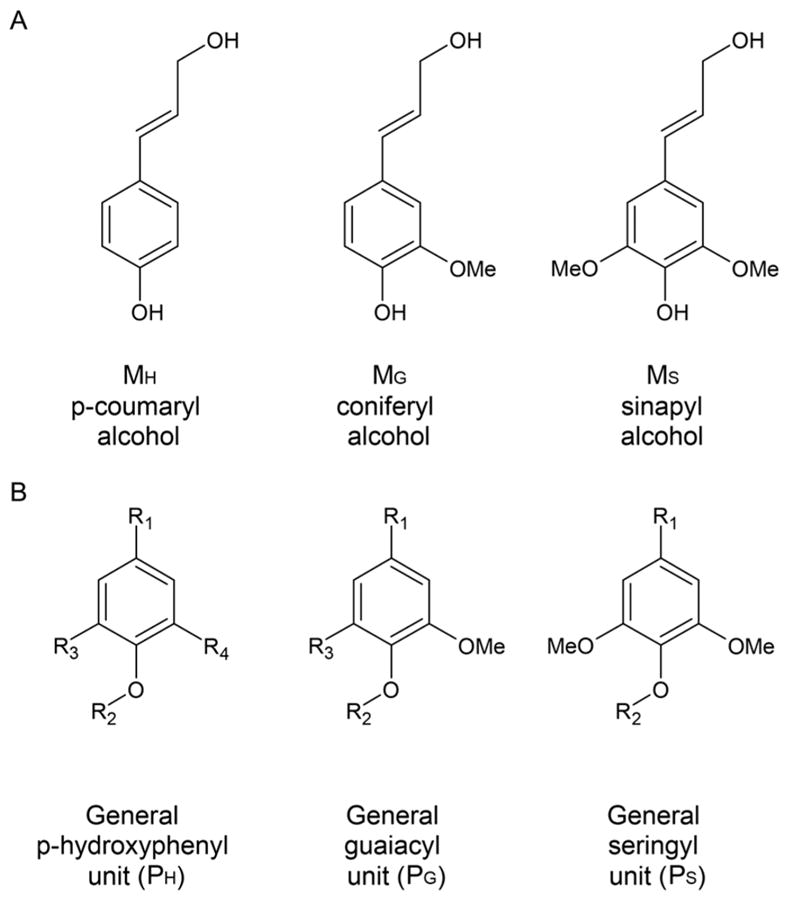
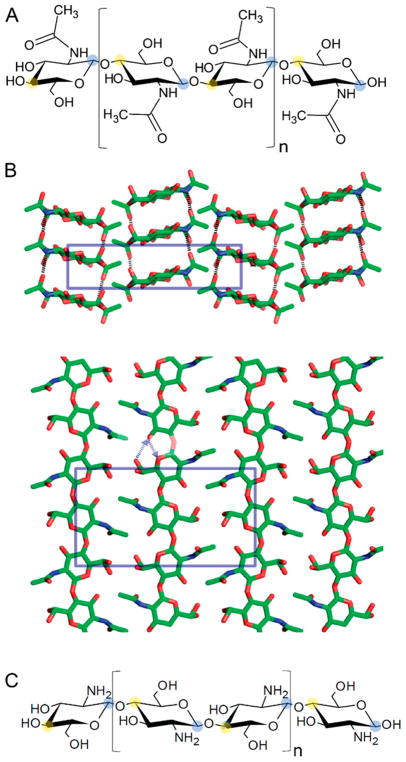
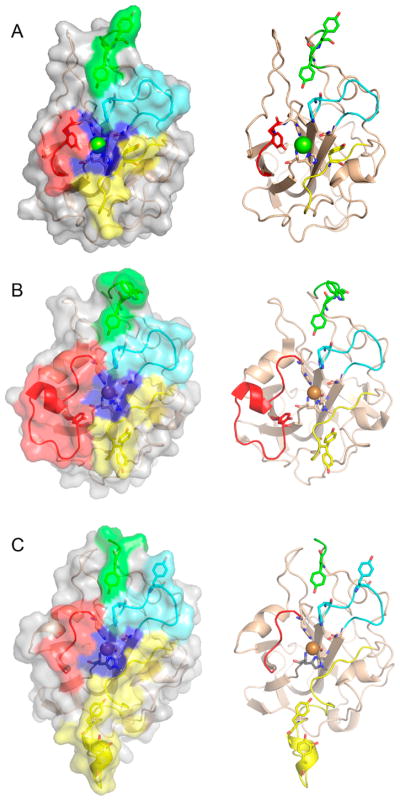
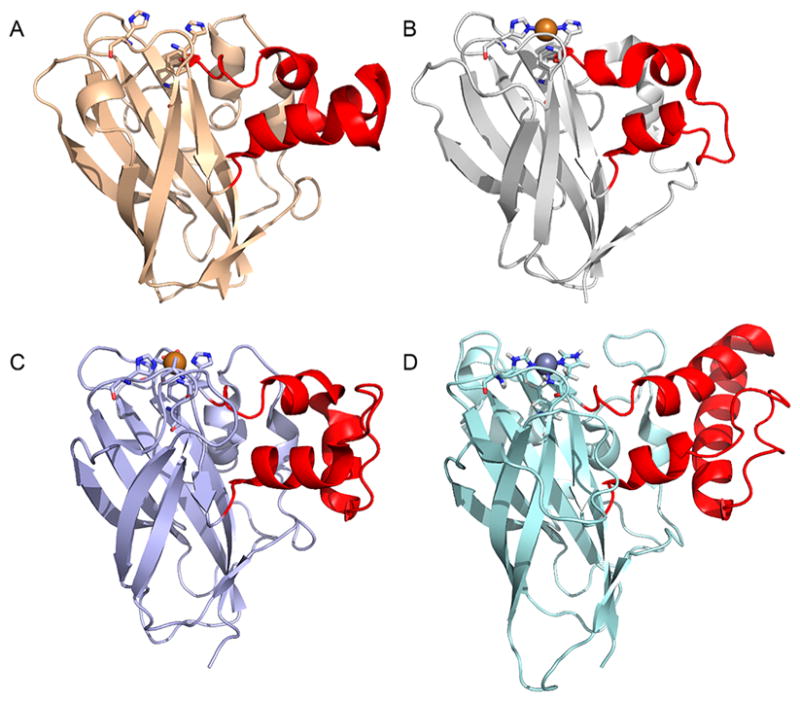
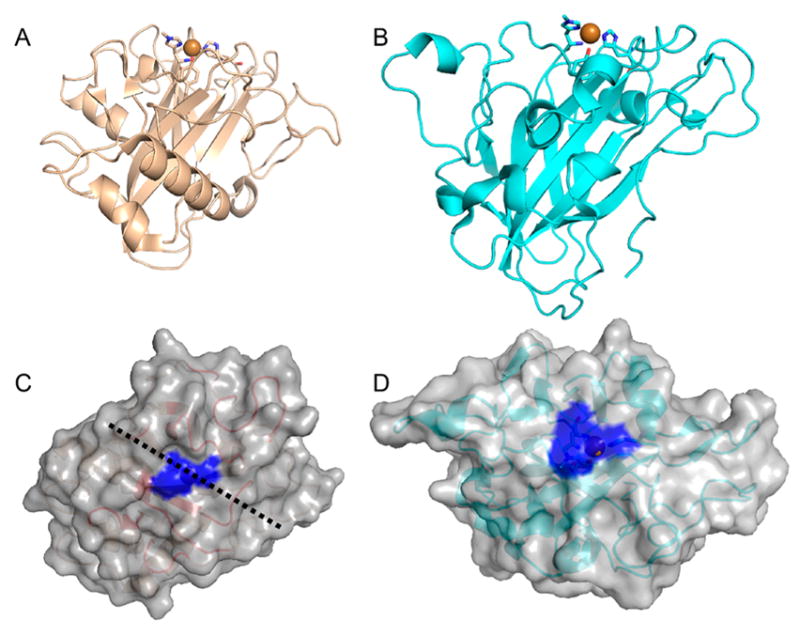
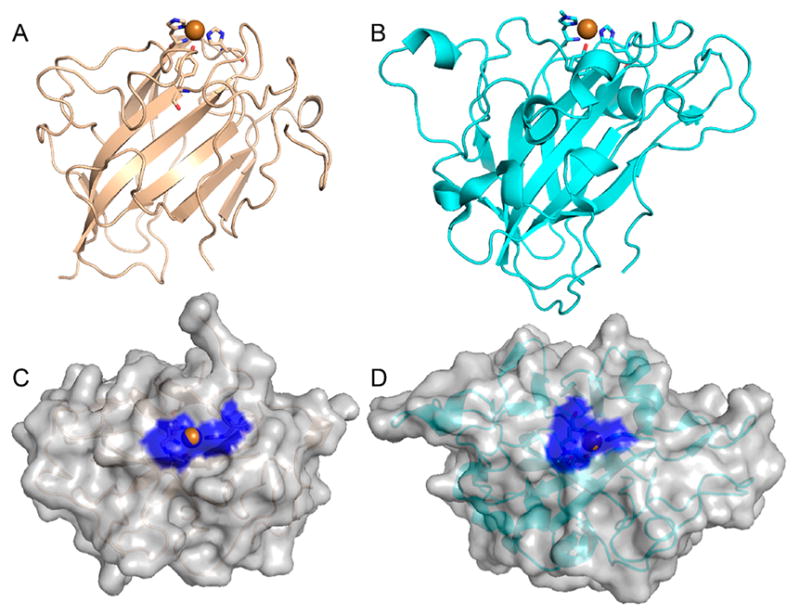
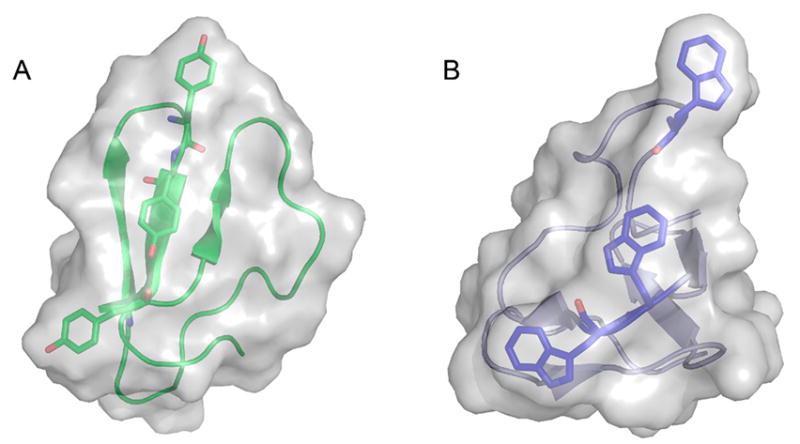
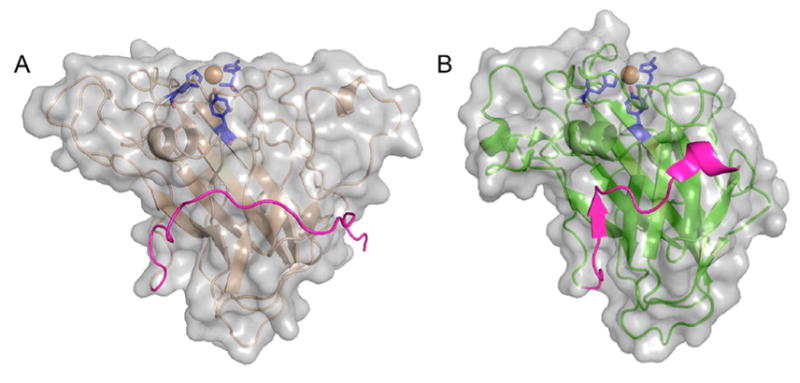
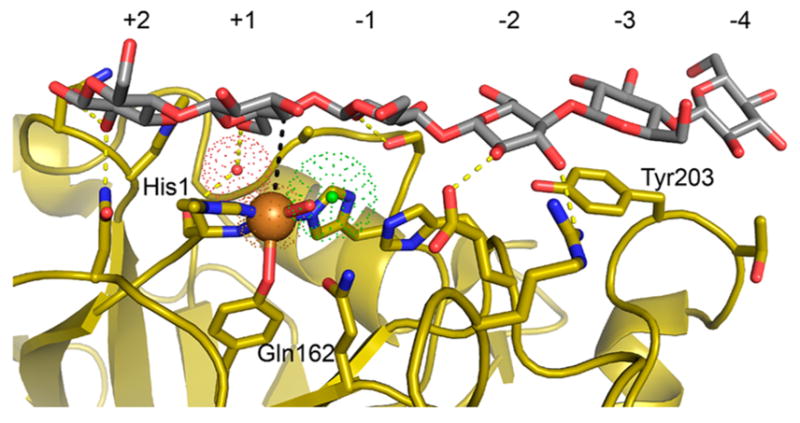
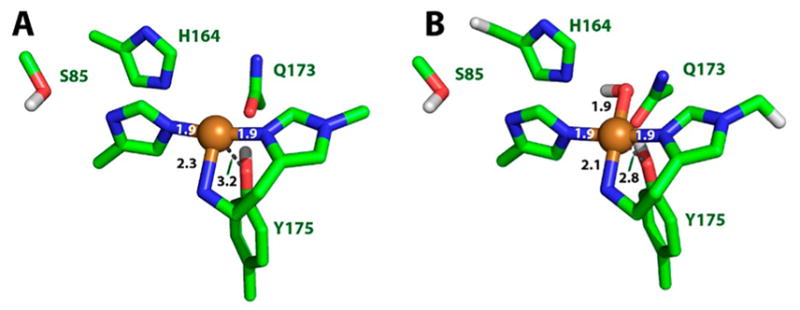
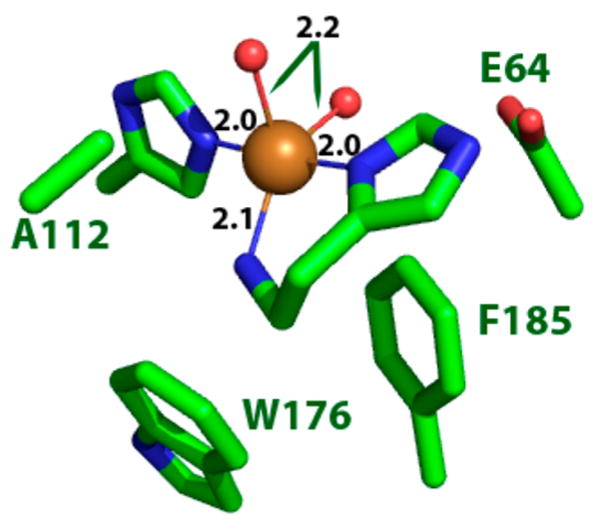
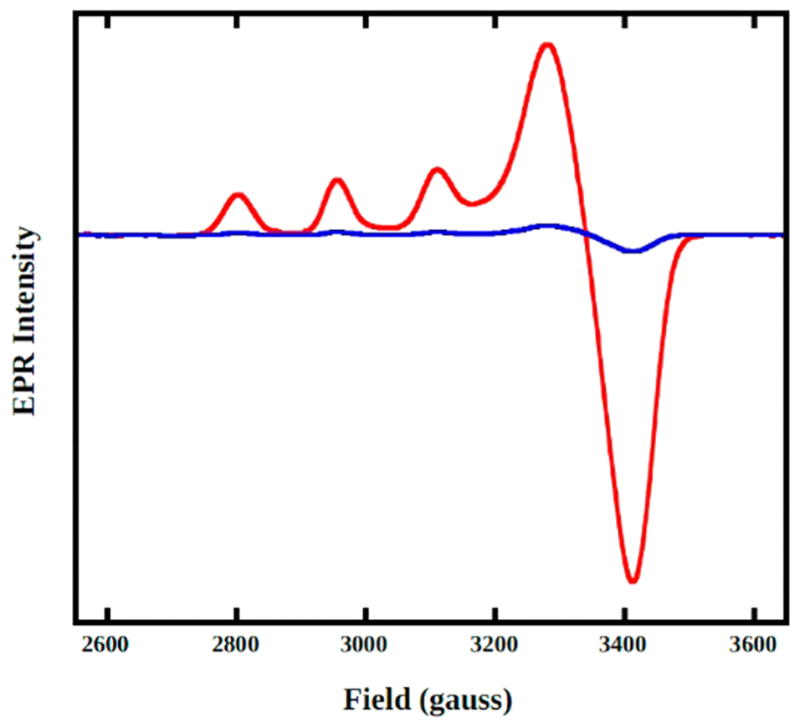
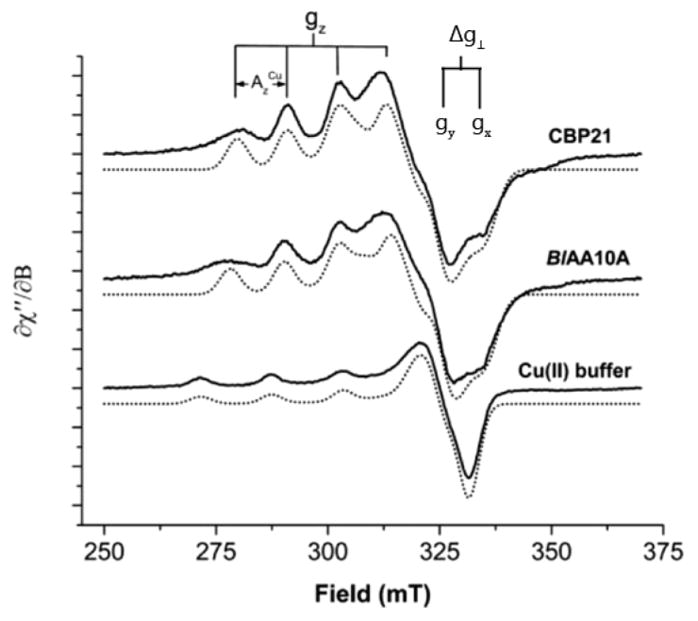
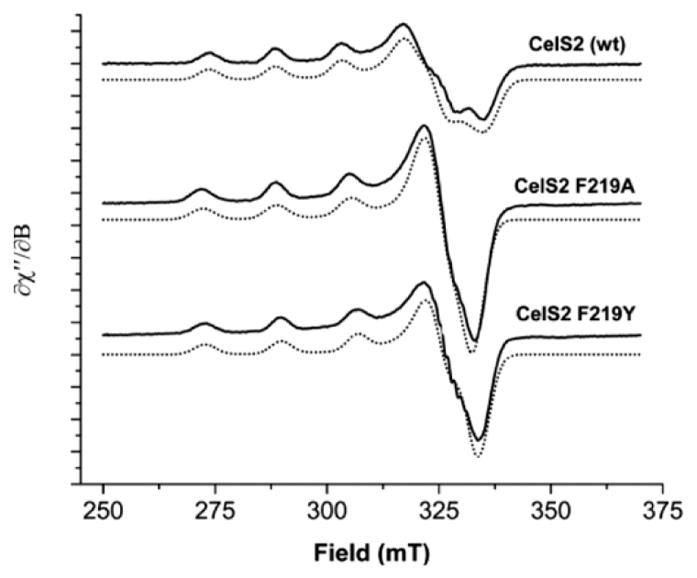
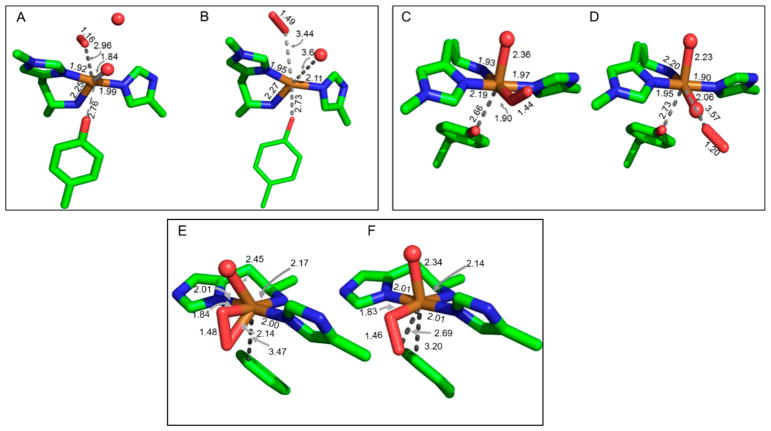
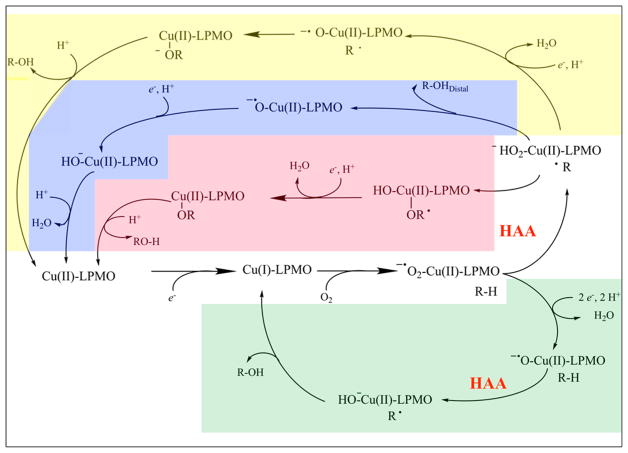
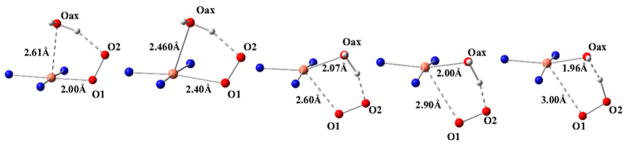
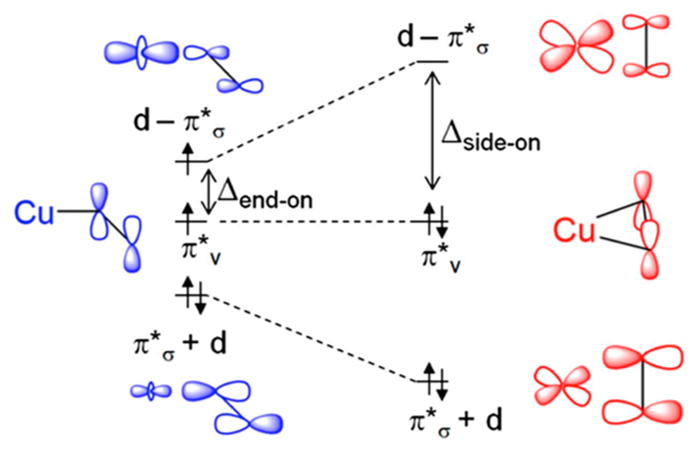
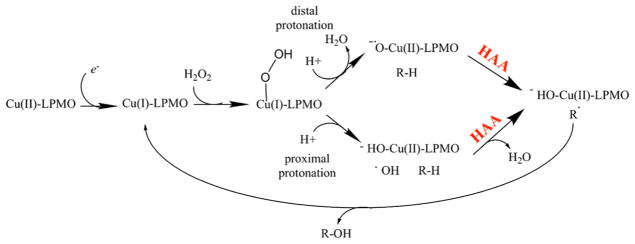
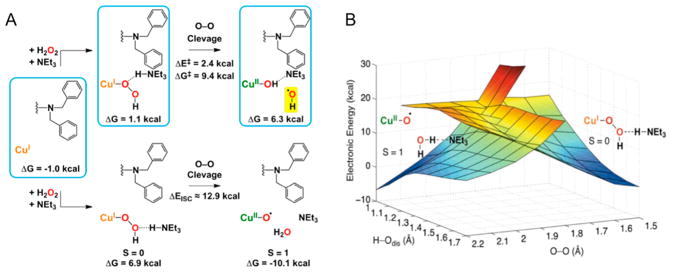
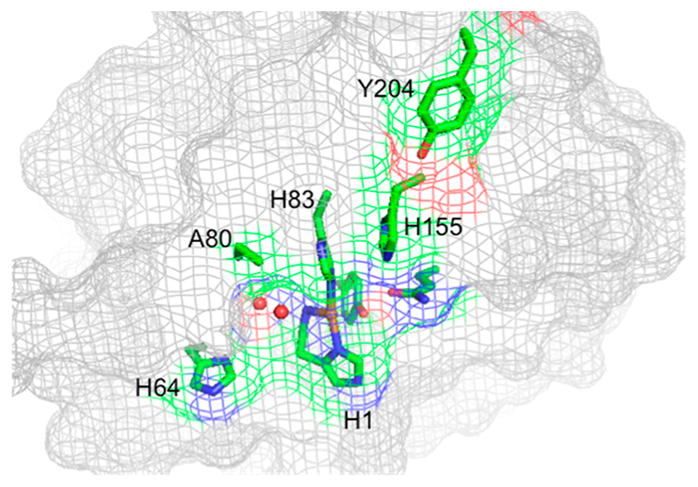
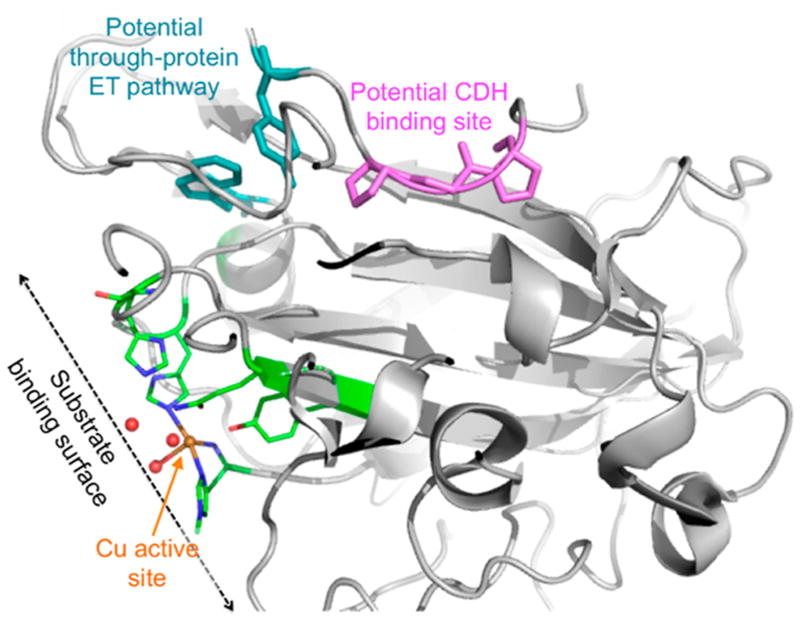
Natural carbohydrate polymers such as starch, cellulose, and chitin provide renewable alternatives to fossil fuels as a source for fuels and materials. As such, there is considerable interest in their conversion for industrial purposes, which is evidenced by the established and emerging markets for products derived from these natural polymers. In many cases, this is achieved via industrial processes that use enzymes to break down carbohydrates to monomer sugars. One of the major challenges facing large-scale industrial applications utilizing natural carbohydrate polymers is rooted in the fact that naturally occurring forms of starch, cellulose, and chitin can have tightly packed organizations of polymer chains with low hydration levels, giving rise to crystalline structures that are highly recalcitrant to enzymatic degradation. The topic of this review is oxidative cleavage of carbohydrate polymers by lytic polysaccharide mono-oxygenases (LPMOs). LPMOs are copper-dependent enzymes (EC 1.14.99.53-56) that, with glycoside hydrolases, participate in the degradation of recalcitrant carbohydrate polymers. Their activity and structural underpinnings provide insights into biological mechanisms of polysaccharide degradation.
天然碳水化合物聚合物,如淀粉、纤维素和几丁质,作为燃料和材料的来源,可以替代化石燃料,提供可再生资源。因此,人们对其转化为工业用途的兴趣相当大,这从这些天然聚合物衍生产品的现有和新兴市场就可以证明。在许多情况下,这是通过使用酶将碳水化合物分解为单体糖的工业过程来实现的。利用天然碳水化合物聚合物的大规模工业应用所面临的主要挑战之一源于这样一个事实,即天然存在的淀粉、纤维素和几丁质形式可能具有聚合物链紧密堆积的组织和低水合水平,导致对酶降解具有高度抗性的结晶结构。本文综述的主题是通过溶菌多糖单加氧酶(LPMO)对碳水化合物聚合物进行氧化裂解。LPMO 是依赖铜的酶(EC 1.14.99.53-56),与糖苷水解酶一起参与难降解碳水化合物聚合物的降解。它们的活性和结构基础为多糖降解的生物学机制提供了深入了解。