Structural Biology Program, Van Andel Research Institute, Grand Rapids, MI, USA.
Department of Molecular Genetics and Microbiology, Stony Brook University, Stony Brook, NY, USA.
Nature. 2018 Oct;562(7727):444-447. doi: 10.1038/s41586-018-0587-z. Epub 2018 Oct 3.
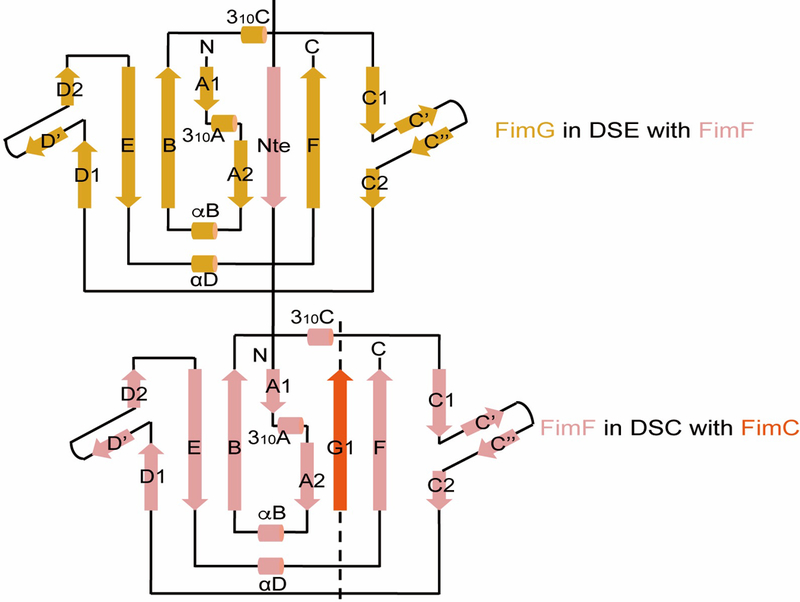
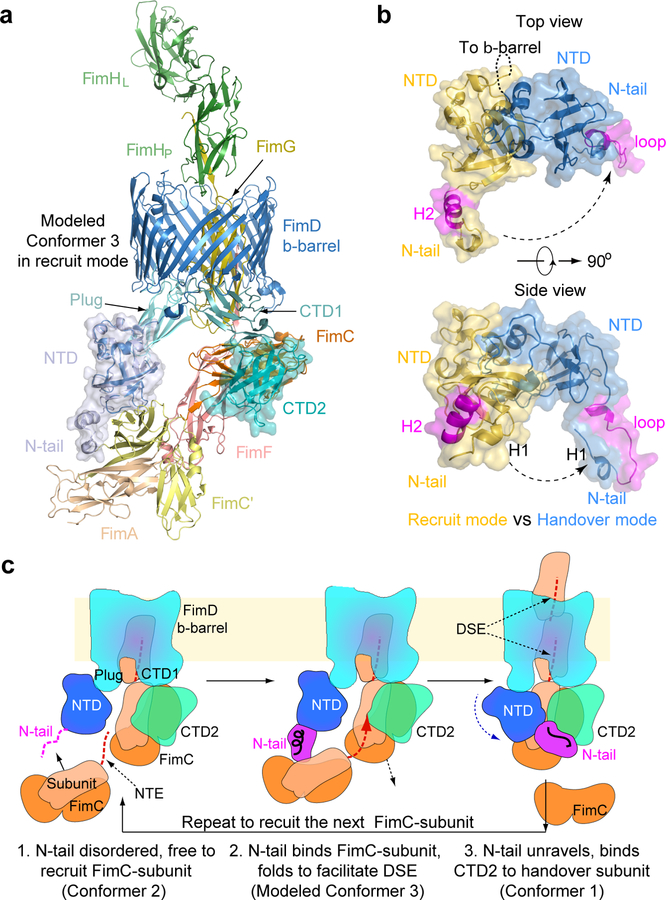
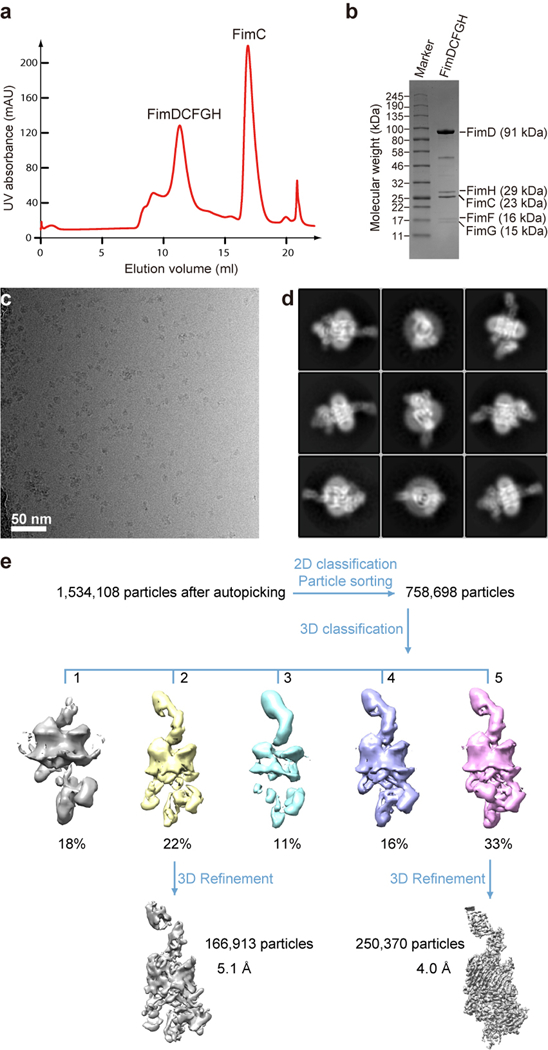
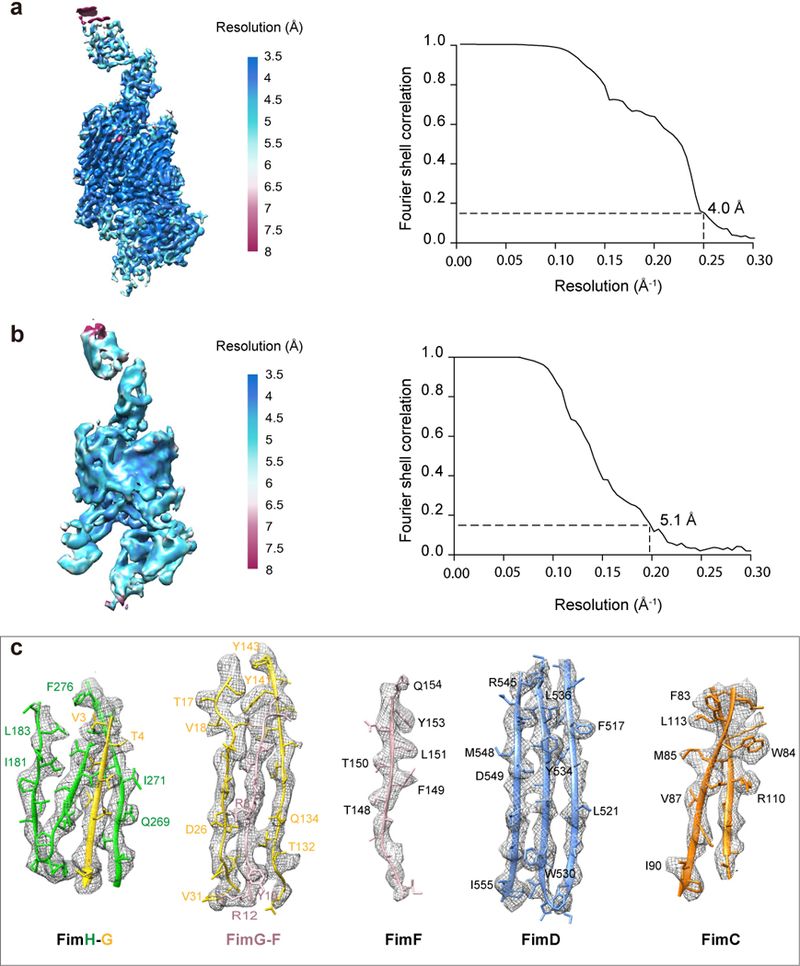
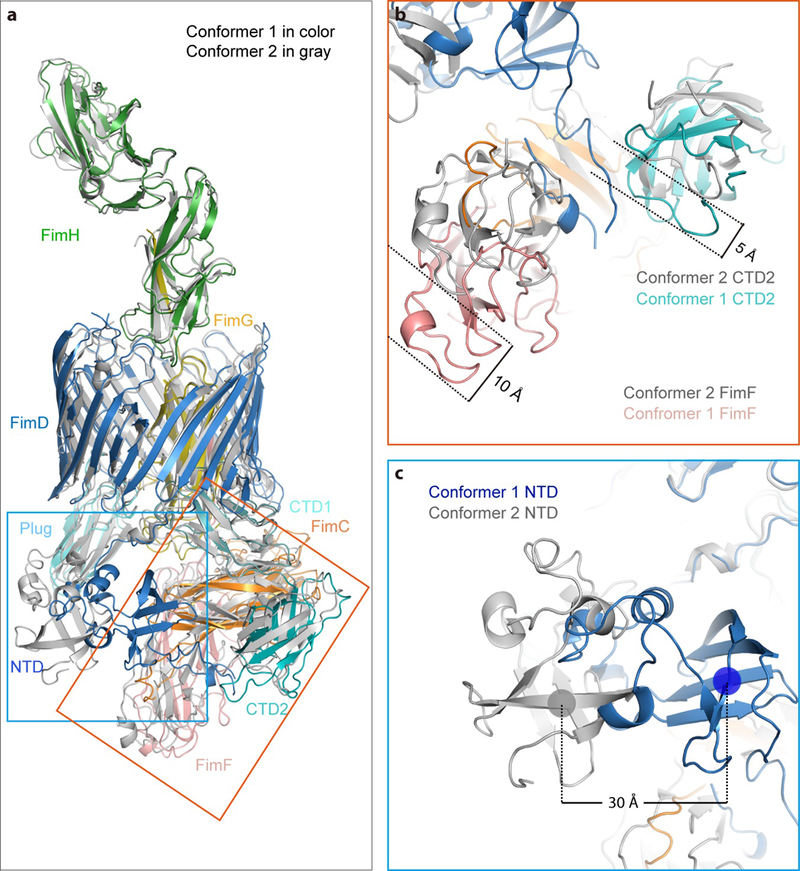
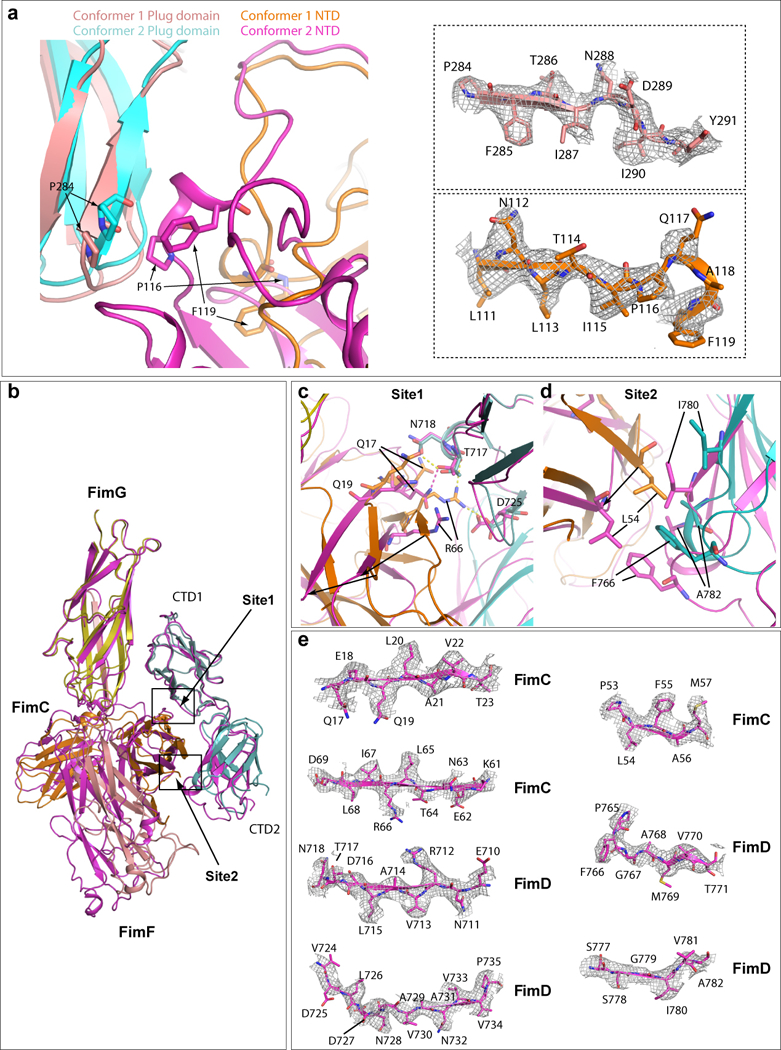
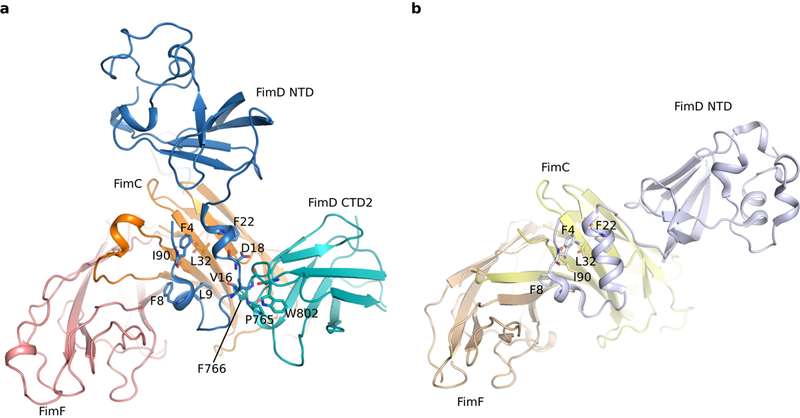
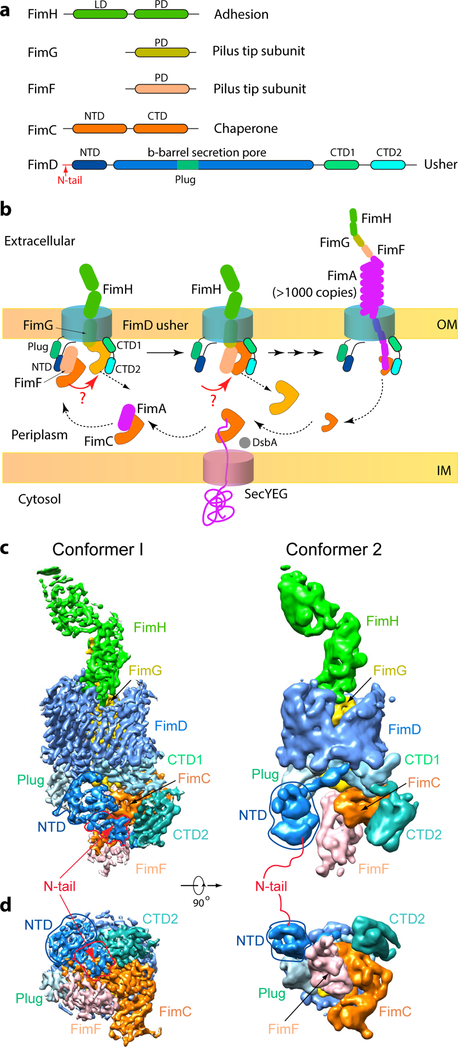
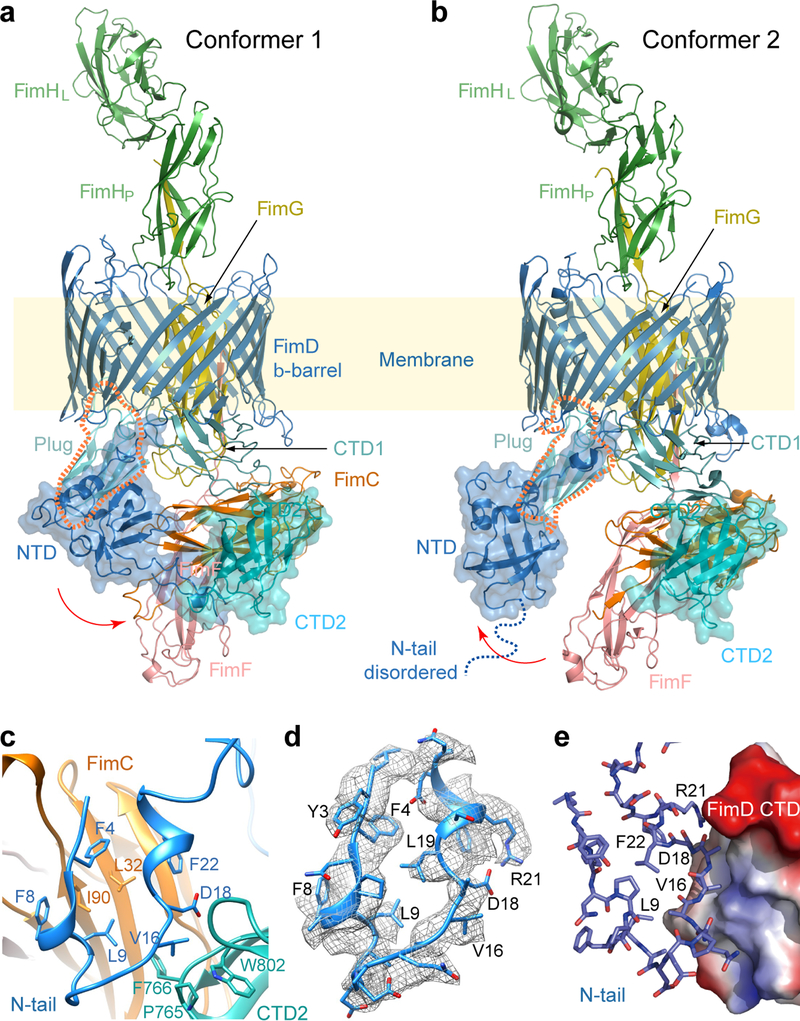
Pathogenic bacteria such as Escherichia coli assemble surface structures termed pili, or fimbriae, to mediate binding to host-cell receptors. Type 1 pili are assembled via the conserved chaperone-usher pathway. The outer-membrane usher FimD recruits pilus subunits bound by the chaperone FimC via the periplasmic N-terminal domain of the usher. Subunit translocation through the β-barrel channel of the usher occurs at the two C-terminal domains (which we label CTD1 and CTD2) of this protein. How the chaperone-subunit complex bound to the N-terminal domain is handed over to the C-terminal domains, as well as the timing of subunit polymerization into the growing pilus, have previously been unclear. Here we use cryo-electron microscopy to capture a pilus assembly intermediate (FimD-FimC-FimF-FimG-FimH) in a conformation in which FimD is in the process of handing over the chaperone-bound end of the growing pilus to the C-terminal domains. In this structure, FimF has already polymerized with FimG, and the N-terminal domain of FimD swings over to bind CTD2; the N-terminal domain maintains contact with FimC-FimF, while at the same time permitting access to the C-terminal domains. FimD has an intrinsically disordered N-terminal tail that precedes the N-terminal domain. This N-terminal tail folds into a helical motif upon recruiting the FimC-subunit complex, but reorganizes into a loop to bind CTD2 during handover. Because both the N-terminal and C-terminal domains of FimD are bound to the end of the growing pilus, the structure further suggests a mechanism for stabilizing the assembly intermediate to prevent the pilus fibre diffusing away during the incorporation of thousands of subunits.
致病细菌如大肠杆菌会组装表面结构,称为菌毛或纤毛,以介导与宿主细胞受体的结合。1 型菌毛通过保守的伴侣-usher 途径组装。外膜 usher FimD 通过 usher 的周质 N 端结构域招募由伴侣 FimC 结合的菌毛亚基。亚基通过 usher 的β桶通道的易位发生在该蛋白的两个 C 端结构域(我们标记为 CTD1 和 CTD2)上。结合到 N 端结构域的伴侣-亚基复合物如何被传递到 C 端结构域,以及亚基聚合到生长菌毛中的时间,以前尚不清楚。在这里,我们使用低温电子显微镜捕获到一个菌毛组装中间体(FimD-FimC-FimF-FimG-FimH),其构象中 FimD 正在将生长菌毛的伴侣结合端传递给 C 端结构域。在这个结构中,FimF 已经与 FimG 聚合,FimD 的 N 端结构域摆动到与 CTD2 结合;N 端结构域与 FimC-FimF 保持接触,同时允许进入 C 端结构域。FimD 具有一个内在无序的 N 端尾巴,位于 N 端结构域之前。该 N 端尾巴在招募 FimC-亚基复合物时折叠成一个螺旋结构,但在传递过程中重新组织成一个环以与 CTD2 结合。由于 FimD 的 N 端和 C 端结构域都与生长菌毛的末端结合,该结构进一步表明了一种稳定组装中间体的机制,以防止在数千个亚基掺入过程中菌毛纤维扩散。