VIB Laboratory for Systems Biology, VIB-KU Leuven Center for Microbiology, Leuven, Belgium.
Departement Microbiële en Moleculaire Systemen (M2S), CMPG Laboratory of Genetics and Genomics, Leuven, Belgium.
Elife. 2018 Oct 9;7:e39234. doi: 10.7554/eLife.39234.
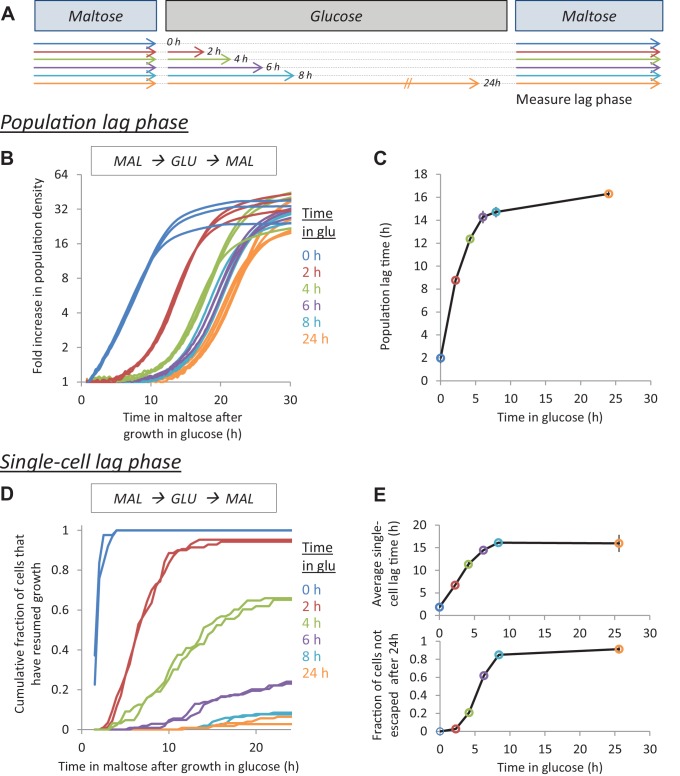
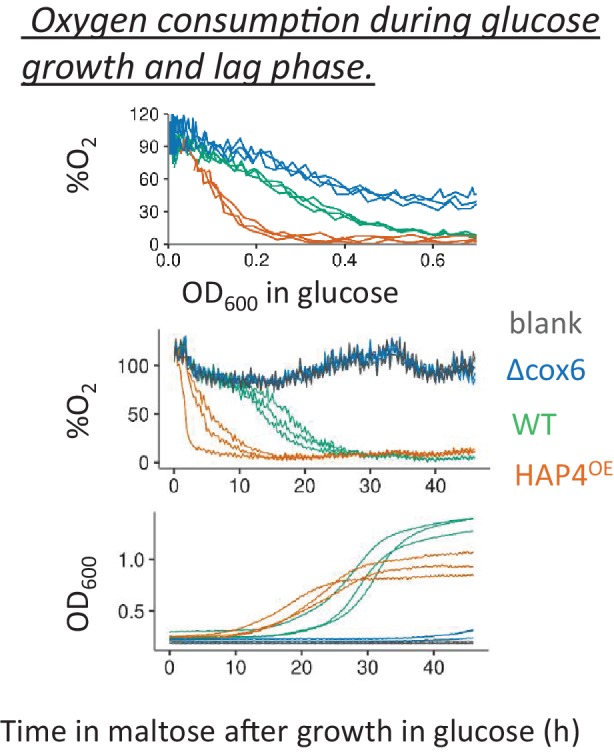
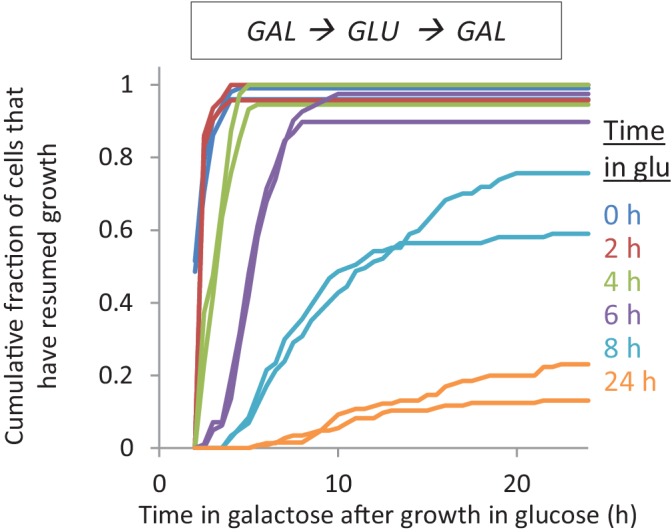
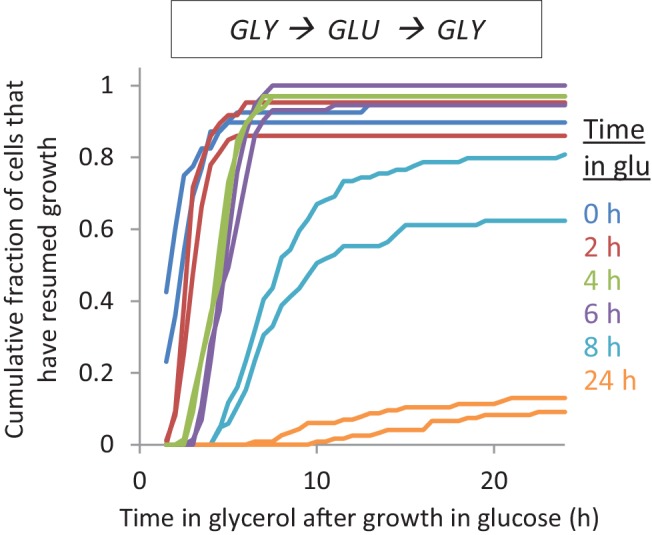
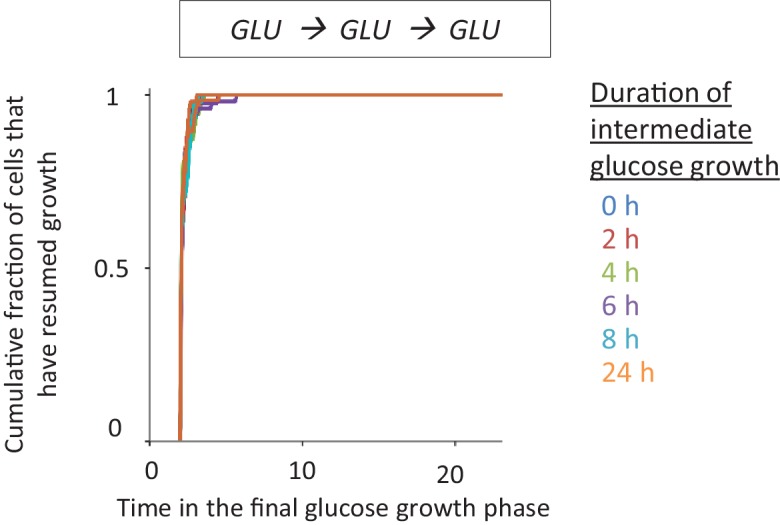
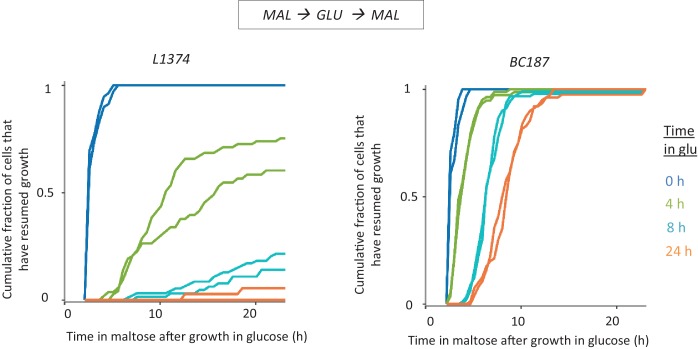
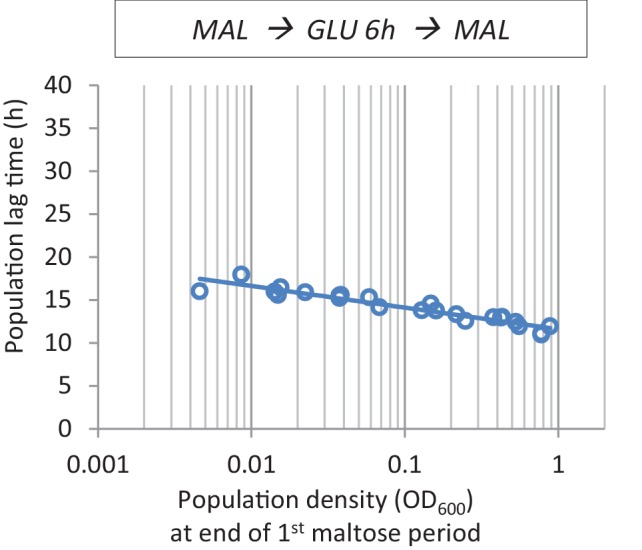
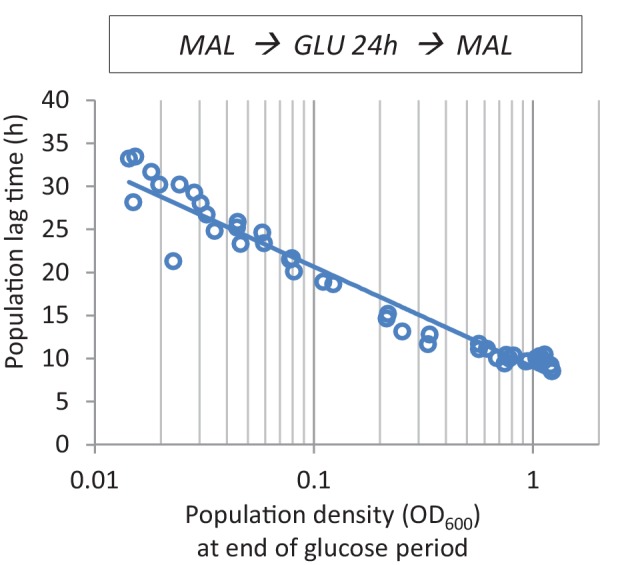
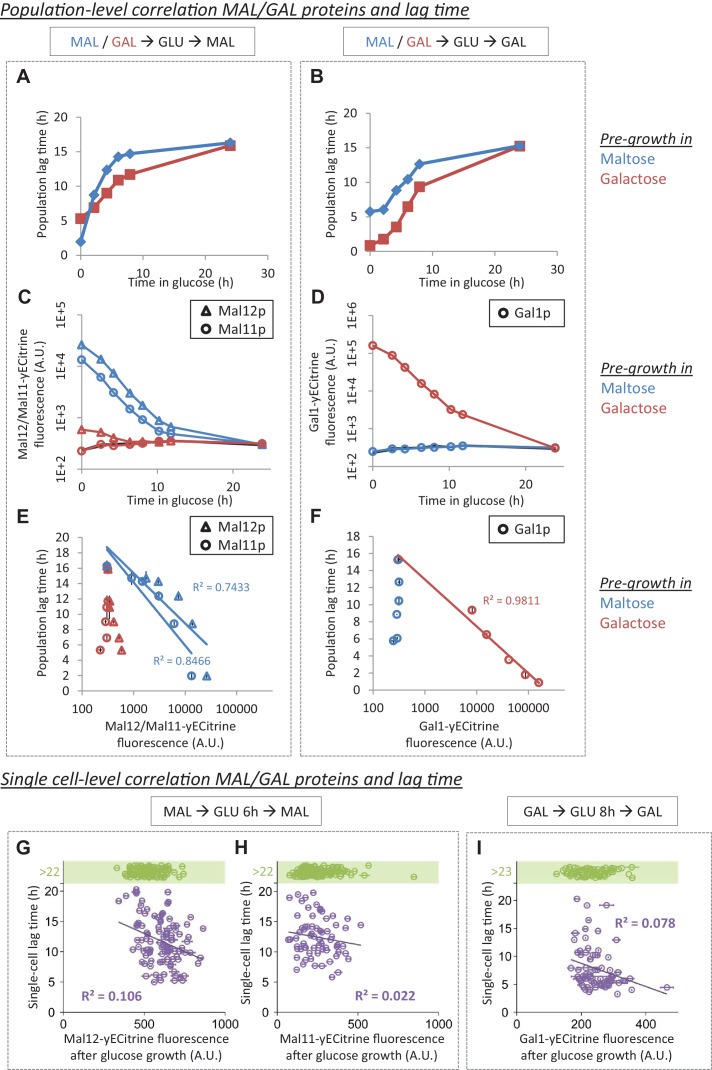
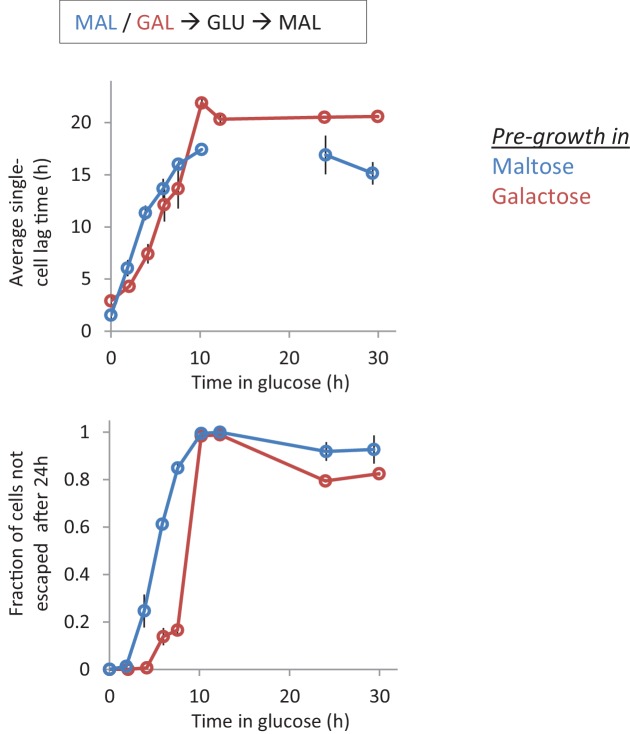
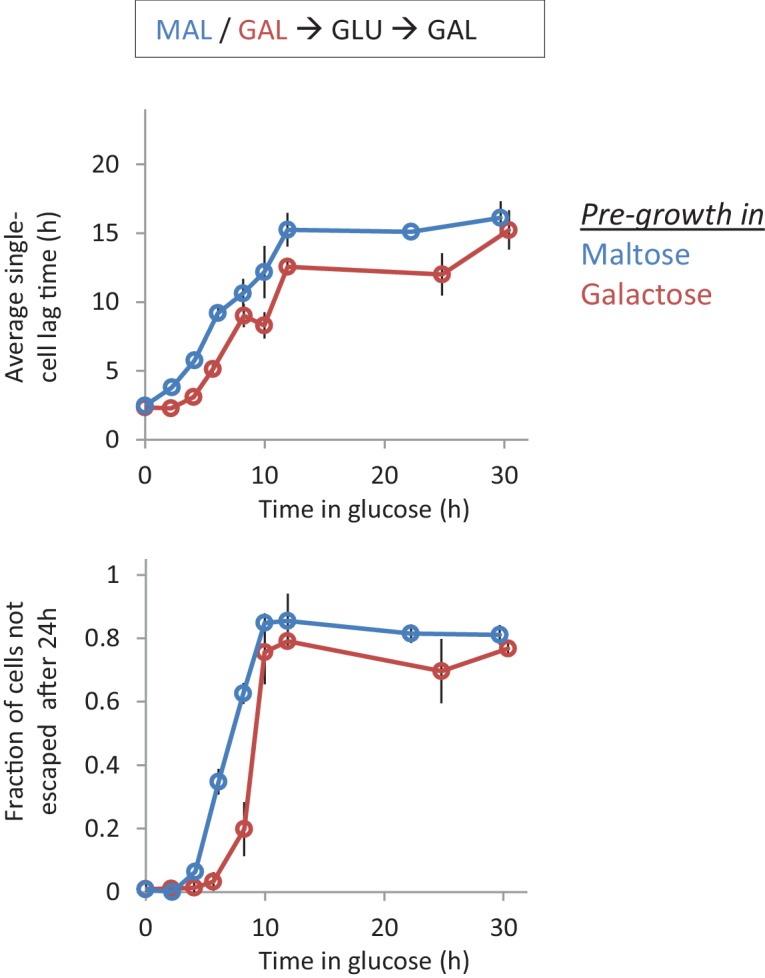
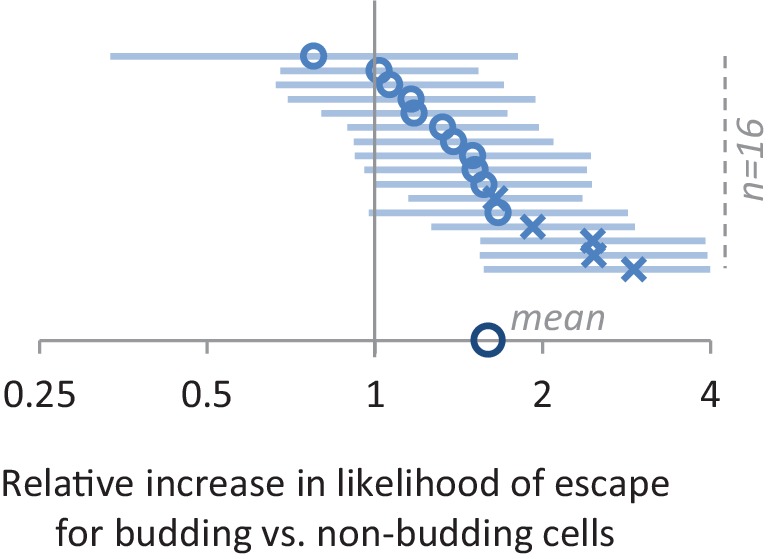
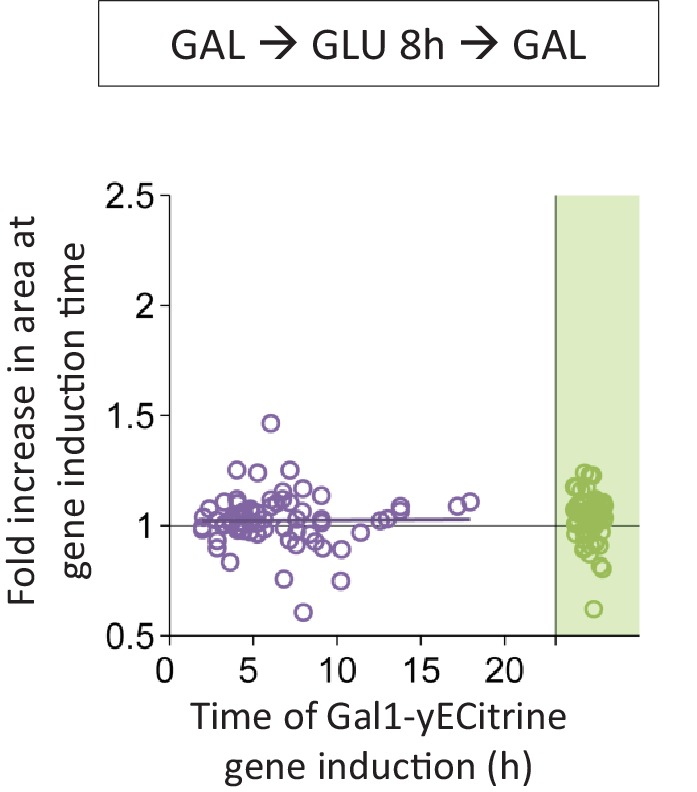
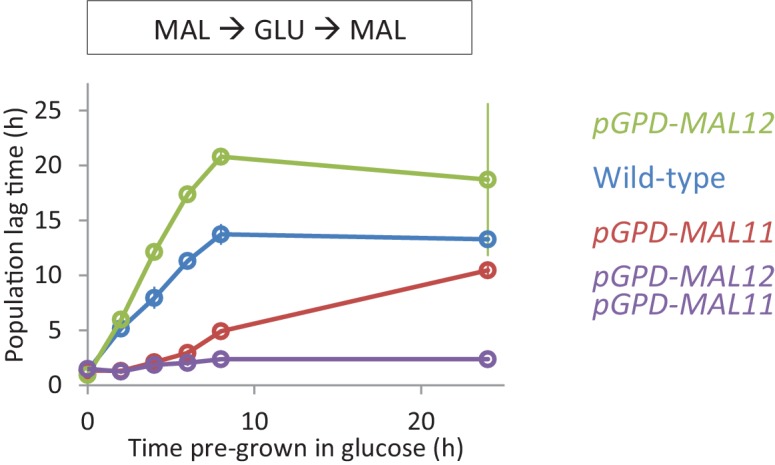
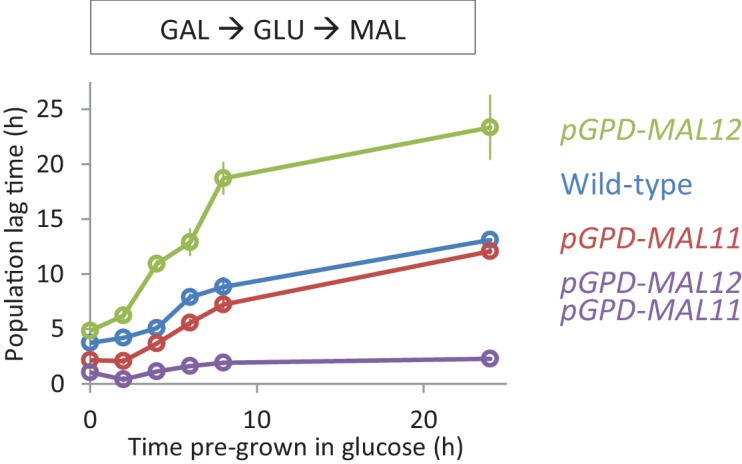
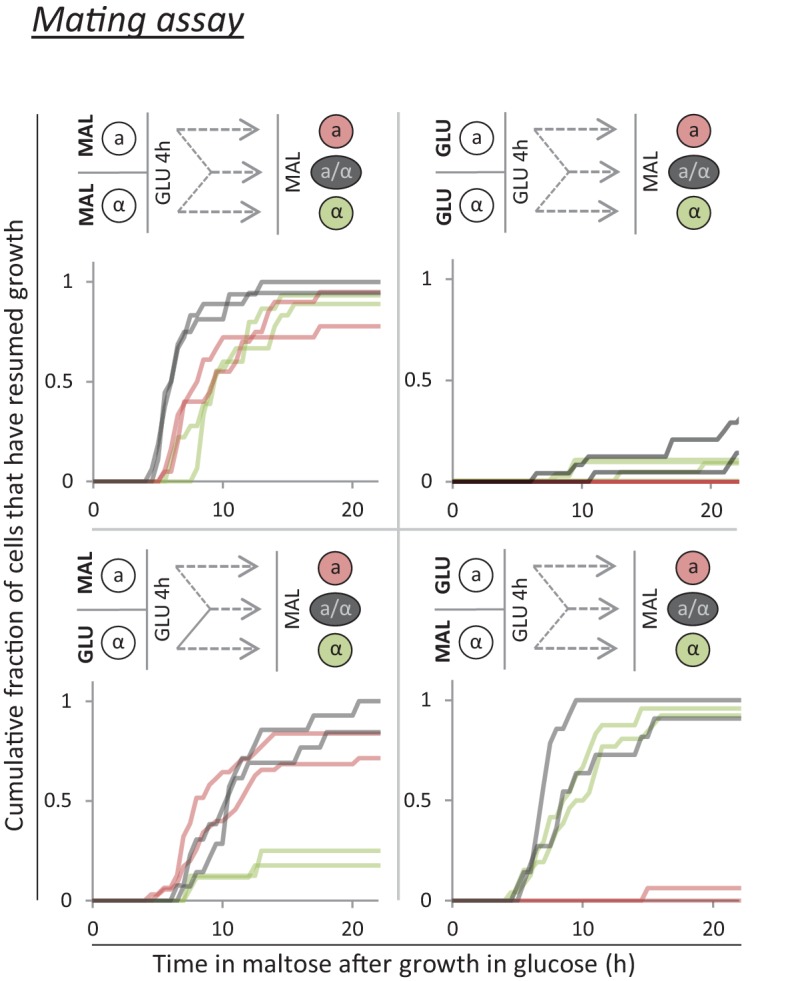
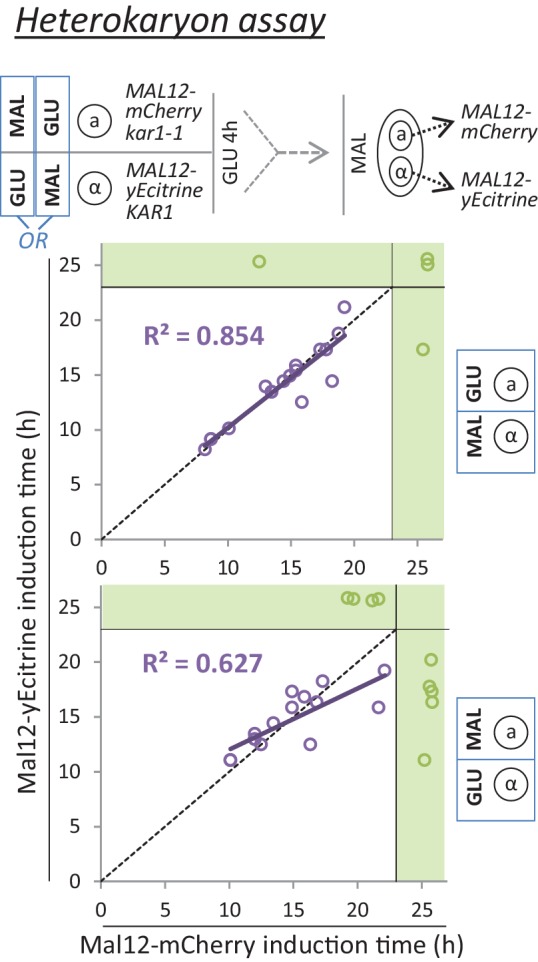
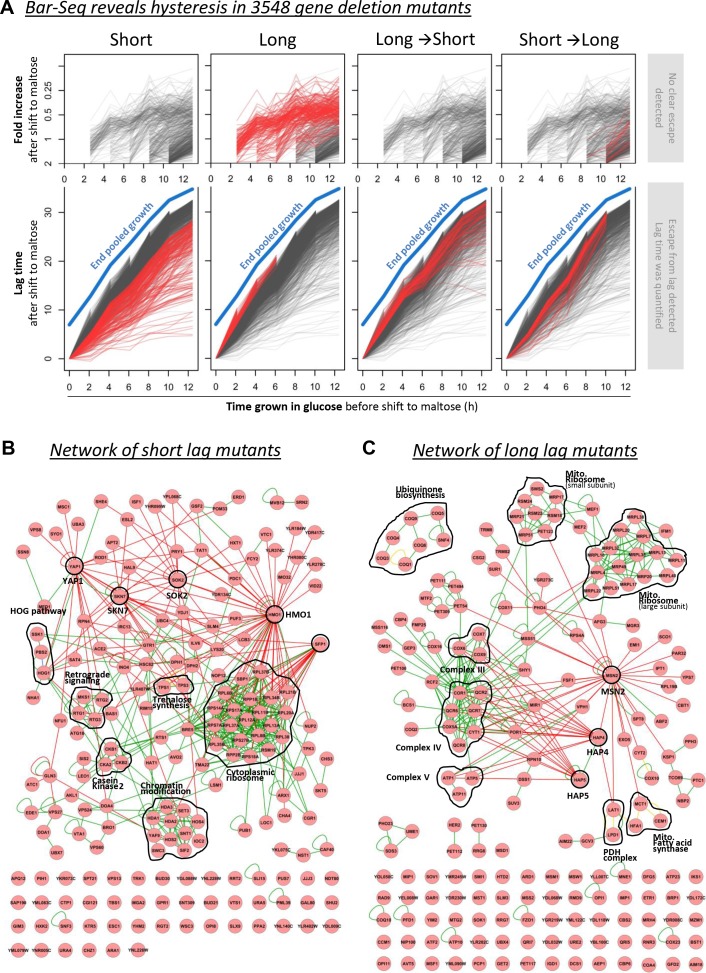
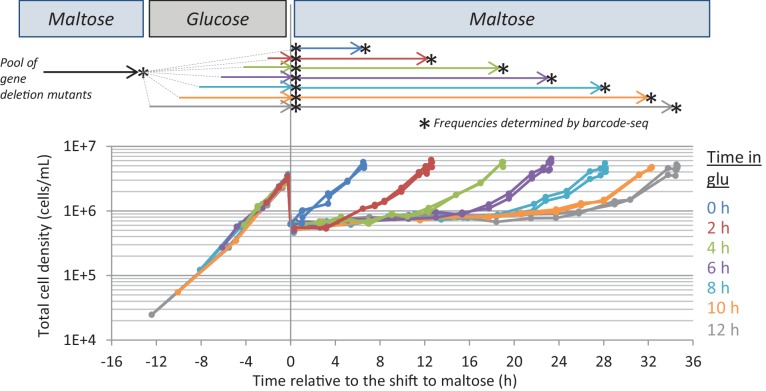
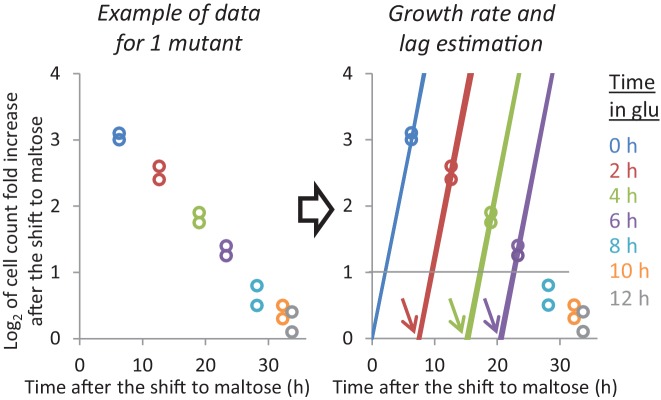
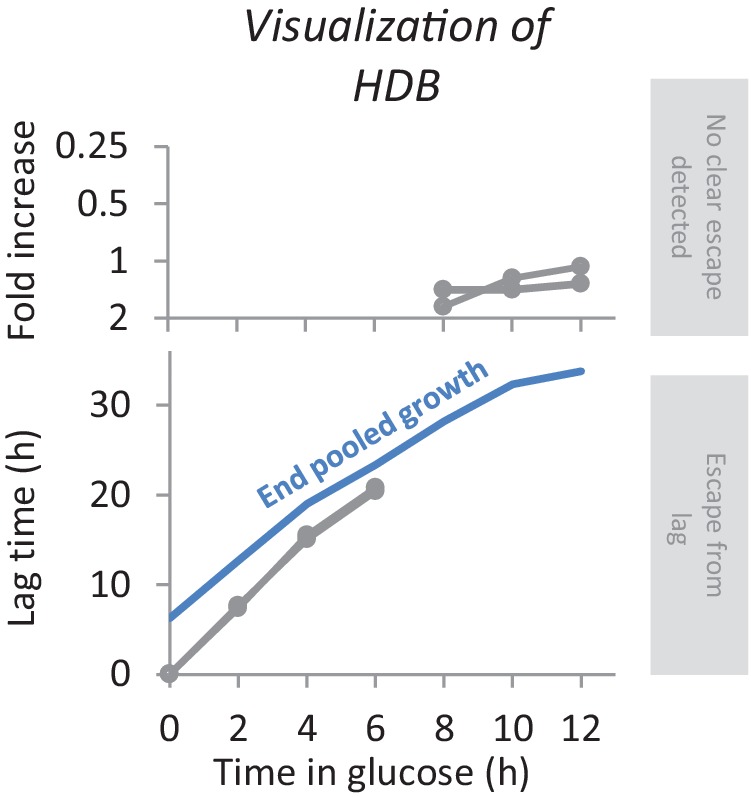
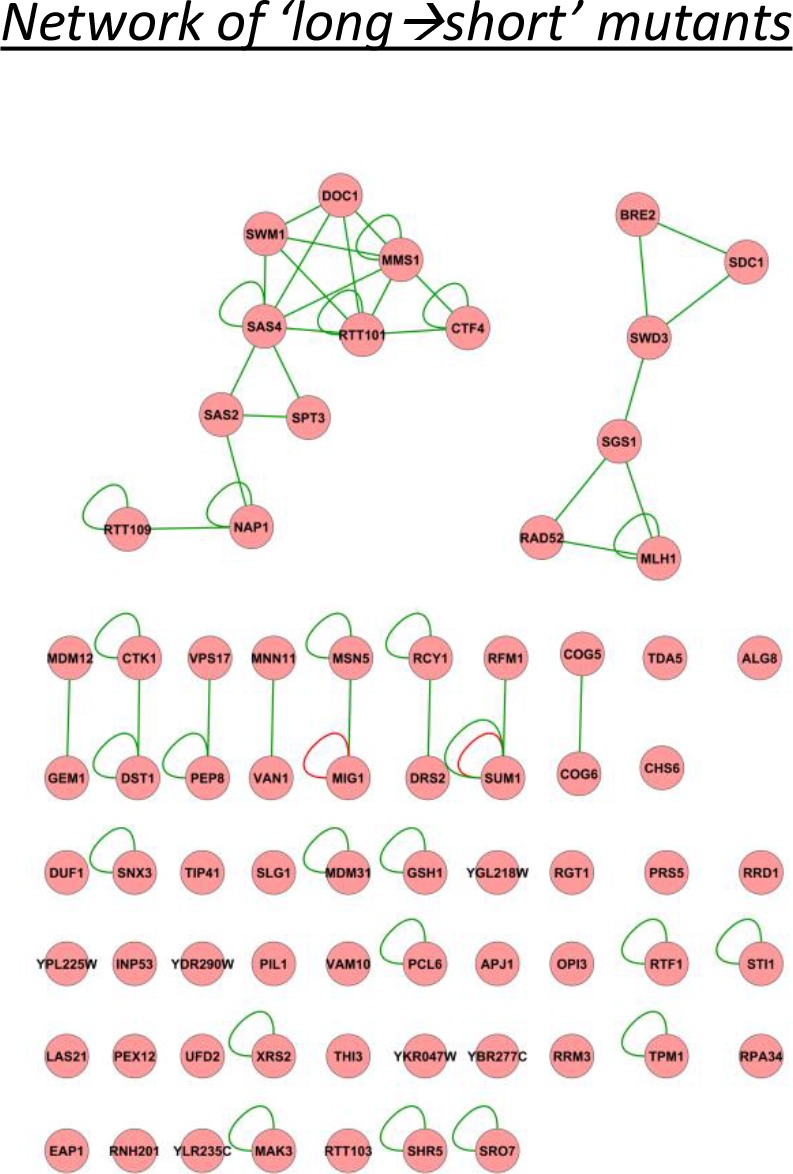
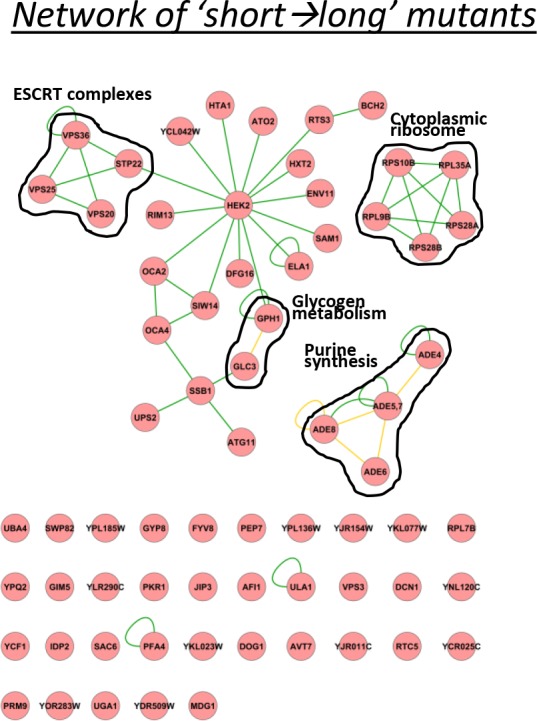
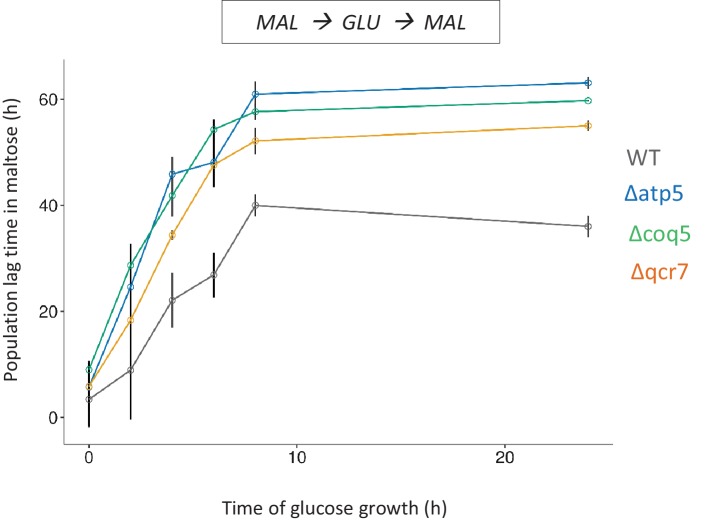
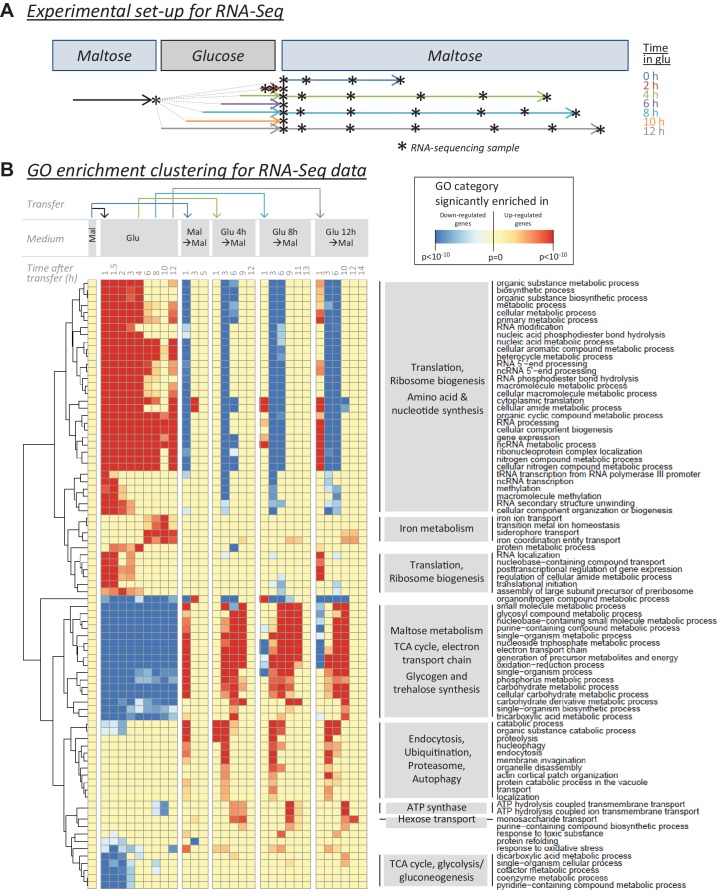
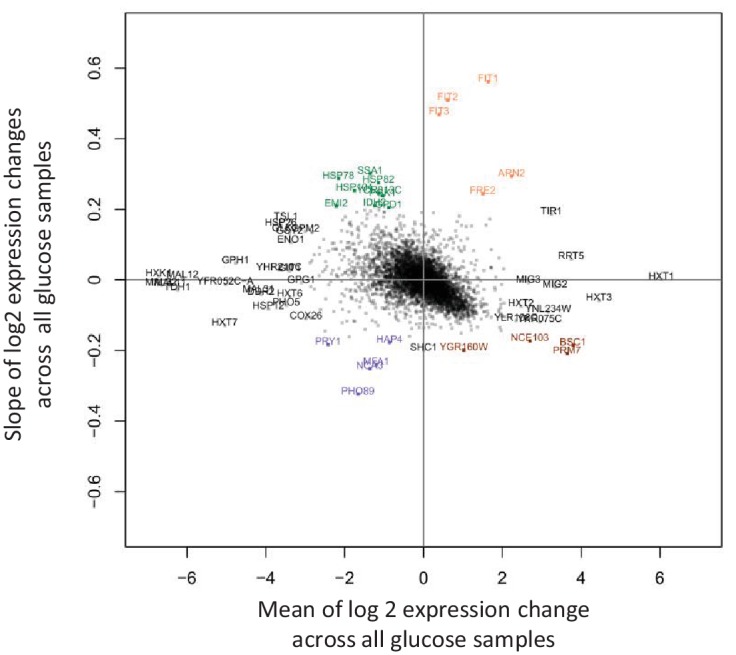
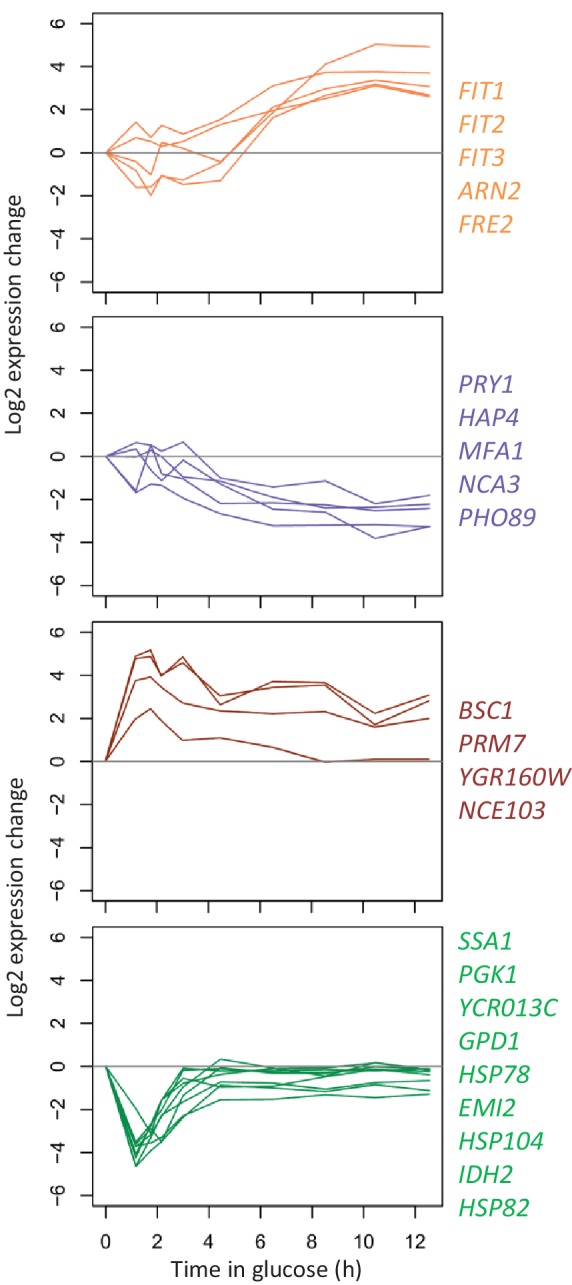
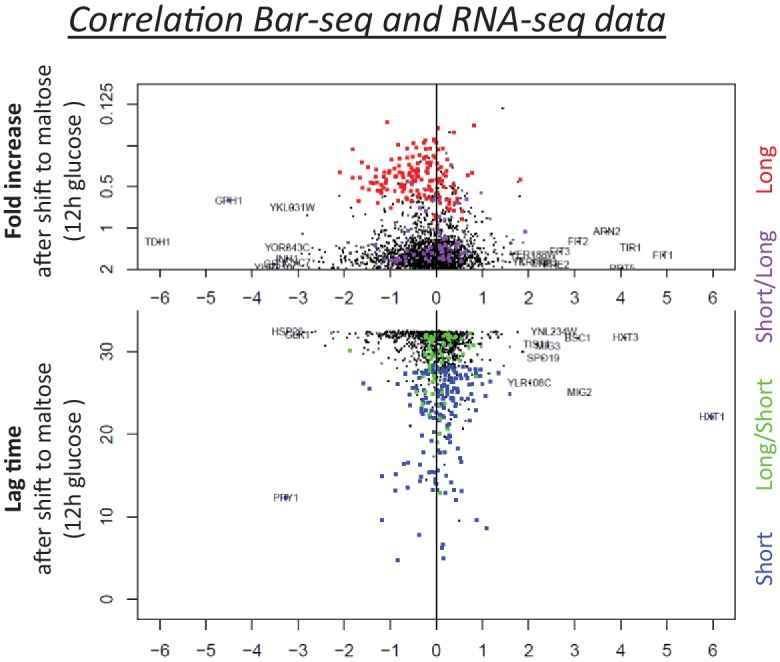
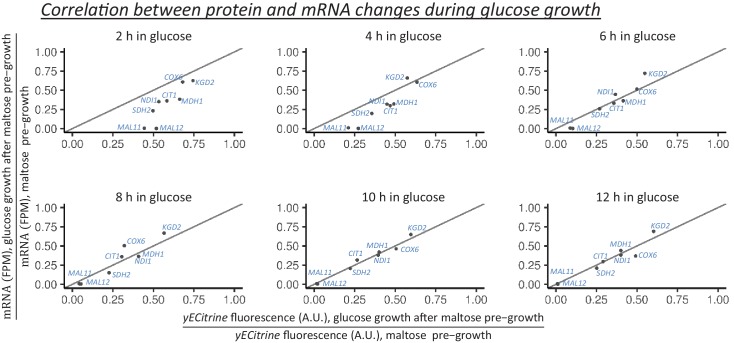
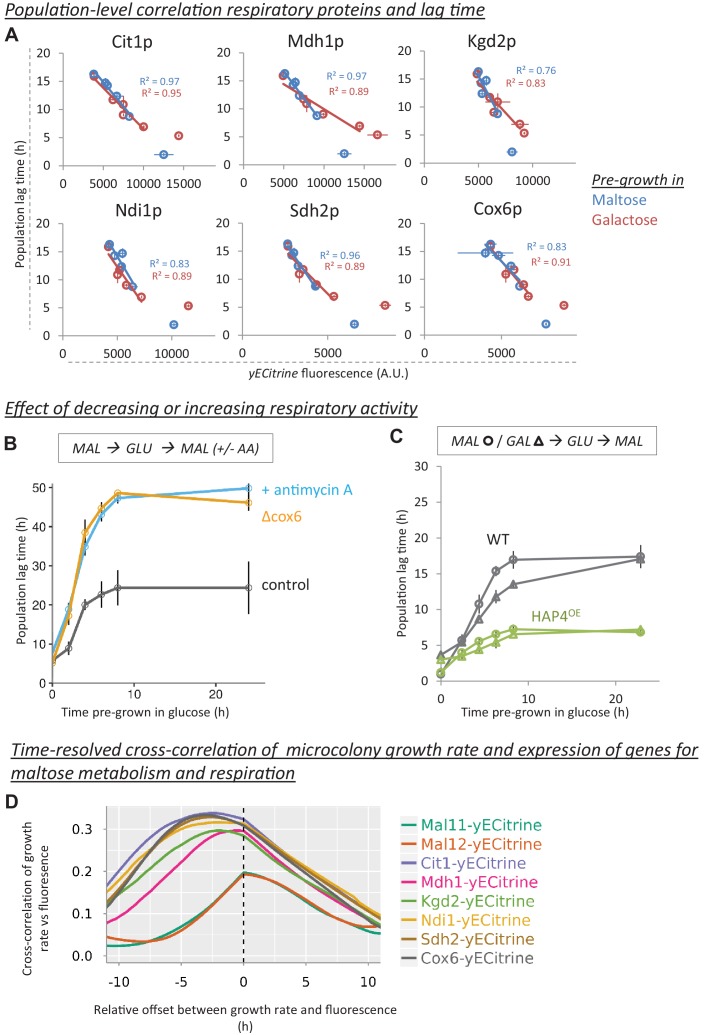
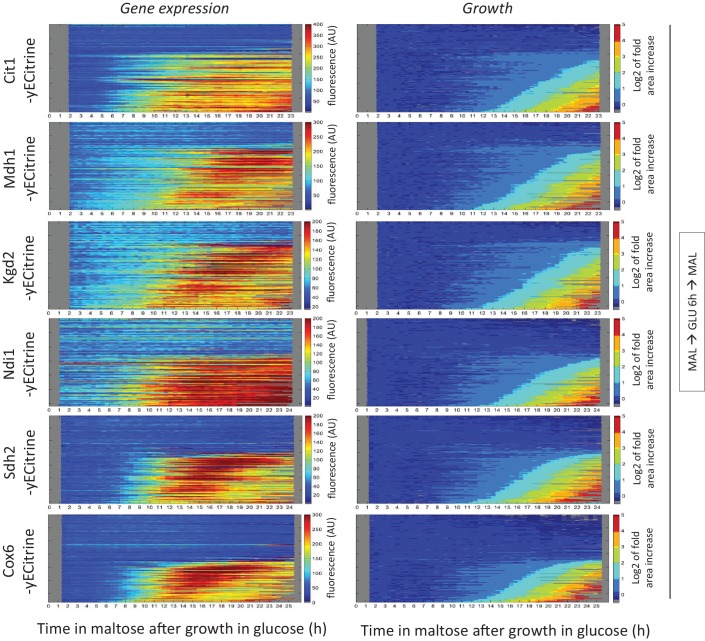
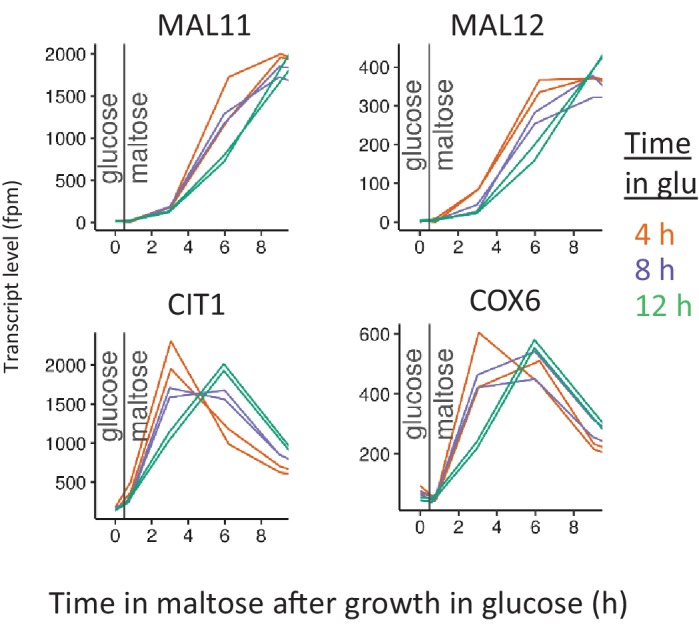
Cells constantly adapt to environmental fluctuations. These physiological changes require time and therefore cause a lag phase during which the cells do not function optimally. Interestingly, past exposure to an environmental condition can shorten the time needed to adapt when the condition re-occurs, even in daughter cells that never directly encountered the initial condition. Here, we use the molecular toolbox of to systematically unravel the molecular mechanism underlying such history-dependent behavior in transitions between glucose and maltose. In contrast to previous hypotheses, the behavior does not depend on persistence of proteins involved in metabolism of a specific sugar. Instead, presence of glucose induces a gradual decline in the cells' ability to activate respiration, which is needed to metabolize alternative carbon sources. These results reveal how trans-generational transitions in central carbon metabolism generate history-dependent behavior in yeast, and provide a mechanistic framework for similar phenomena in other cell types.
细胞不断适应环境波动。这些生理变化需要时间,因此在细胞不能最佳工作的迟滞期内会产生滞后。有趣的是,过去暴露于环境条件下会缩短再次出现该条件时所需的适应时间,即使是从未直接遇到初始条件的子细胞也是如此。在这里,我们使用的分子工具包来系统地揭示在葡萄糖和麦芽糖之间的转换过程中,这种历史依赖性行为背后的分子机制。与之前的假设相反,这种行为不依赖于参与特定糖代谢的蛋白质的持续存在。相反,葡萄糖的存在会逐渐降低细胞激活呼吸的能力,而呼吸是代谢替代碳源所必需的。这些结果揭示了中央碳代谢的跨代转变如何在酵母中产生历史依赖性行为,并为其他细胞类型中的类似现象提供了一个机械框架。