Aubrey Karin R, Supplisson Stéphane
Institut de Biologie de l'Ecole Normale Supérieure (IBENS), Ecole Normale Supérieure, CNRS, INSERM, PSL Université Paris Paris, France.
Neurobiology of Pain Laboratory, Kolling Institute, Royal North Shore Hospital St. Leonards, NSW, Australia.
Front Synaptic Neurosci. 2018 Nov 6;10:40. doi: 10.3389/fnsyn.2018.00040. eCollection 2018.
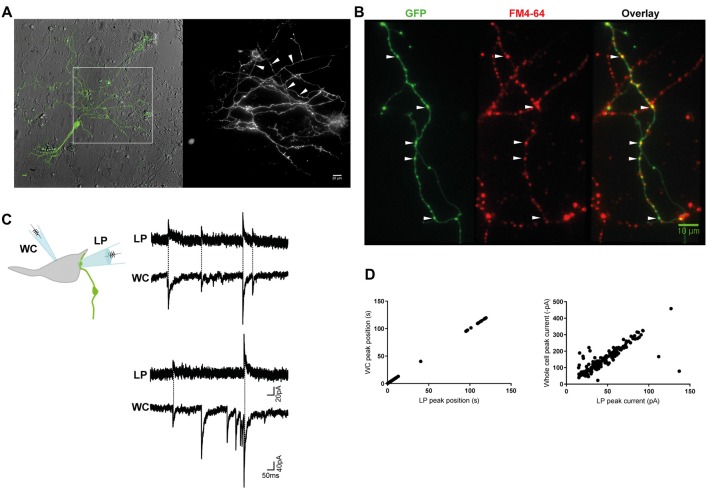
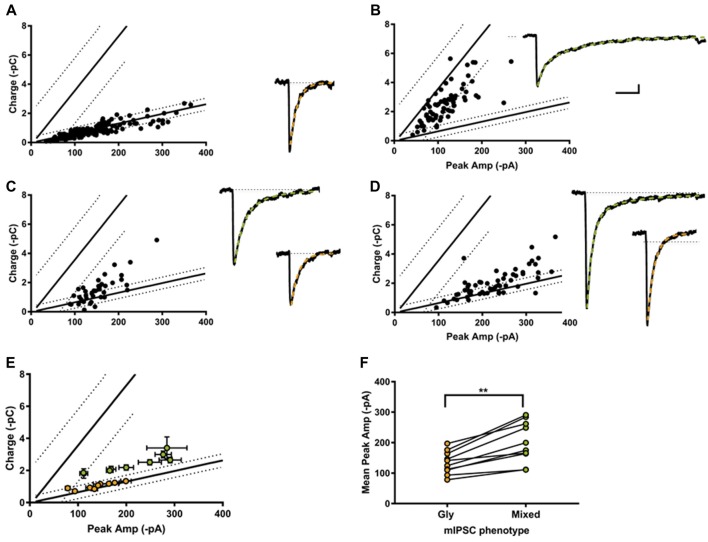
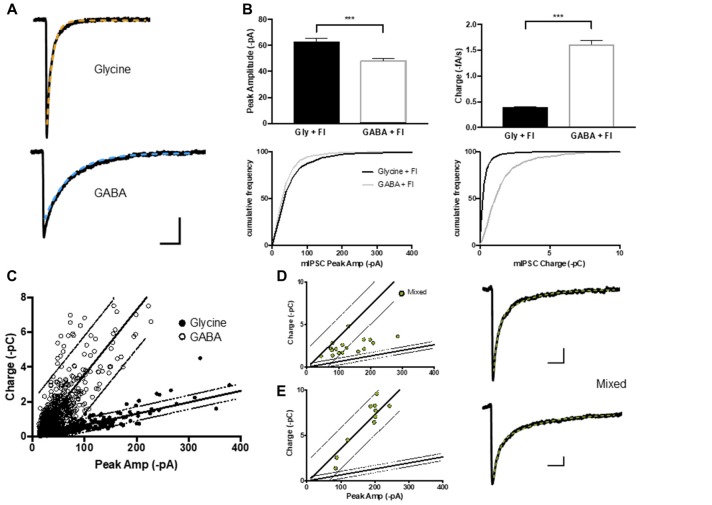
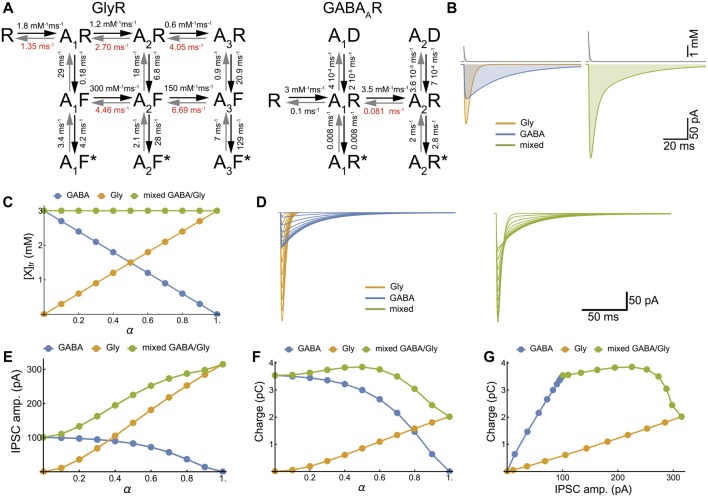
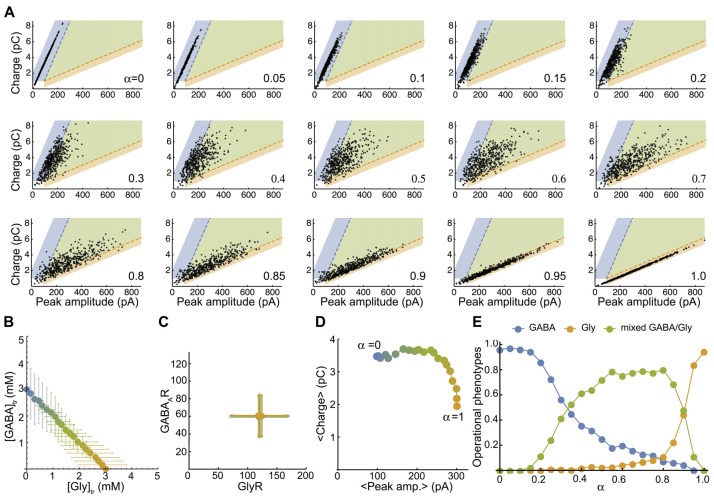
The corelease of several neurotransmitters from a single synaptic vesicle has been observed at many central synapses. Nevertheless, the signaling synergy offered by cotransmission and the mechanisms that maintain the optimal release and detection of neurotransmitters at mixed synapses remain poorly understood, thus limiting our ability to interpret changes in synaptic signaling and identify molecules important for plasticity. In the brainstem and spinal cord, GABA and glycine cotransmission is facilitated by a shared vesicular transporter VIAAT (also named VGAT), and occurs at many immature inhibitory synapses. As sensory and motor networks mature, GABA/glycine cotransmission is generally replaced by either pure glycinergic or GABAergic transmission, and the functional role for the continued corelease of GABA and glycine is unclear. Whether or not, and how, the GABA/glycine content is balanced in VIAAT-expressing vesicles from the same terminal, and how loading variability effects the strength of inhibitory transmission is not known. Here, we use a combination of loose-patch (LP) and whole-cell (WC) electrophysiology in cultured spinal neurons of GlyT2:eGFP mice to sample miniature inhibitory post synaptic currents (mIPSCs) that originate from individual GABA/glycine co-releasing synapses and develop a modeling approach to illustrate the gradual change in mIPSC phenotypes as glycine replaces GABA in vesicles. As a consistent GABA/glycine balance is predicted if VIAAT has access to both amino-acids, we test whether vesicle exocytosis from a single terminal evokes a homogeneous population of mixed mIPSCs. We recorded mIPSCs from 18 individual synapses and detected glycine-only mIPSCs in 4/18 synapses sampled. The rest (14/18) were co-releasing synapses that had a significant proportion of mixed GABA/glycine mIPSCs with a characteristic biphasic decay. The majority (9/14) of co-releasing synapses did not have a homogenous phenotype, but instead signaled with a combination of mixed and pure mIPSCs, suggesting that there is variability in the loading and/or storage of GABA and glycine at the level of individual vesicles. Our modeling predicts that when glycine replaces GABA in synaptic vesicles, the redistribution between the peak amplitude and charge transfer of mIPSCs acts to maintain the strength of inhibition while increasing the temporal precision of signaling.
在许多中枢突触中已观察到单个突触小泡可共同释放多种神经递质。然而,共传递所提供的信号协同作用以及在混合突触中维持神经递质最佳释放和检测的机制仍知之甚少,这限制了我们解释突触信号变化以及识别对可塑性重要分子的能力。在脑干和脊髓中,GABA和甘氨酸的共传递由共同的囊泡转运体VIAAT(也称为VGAT)促进,并发生在许多未成熟的抑制性突触中。随着感觉和运动网络的成熟,GABA/甘氨酸共传递通常会被纯甘氨酸能或GABA能传递所取代,而GABA和甘氨酸持续共释放的功能作用尚不清楚。来自同一终末的表达VIAAT的囊泡中GABA/甘氨酸含量是否平衡以及如何平衡,以及装载变异性如何影响抑制性传递的强度尚不清楚。在这里,我们在GlyT2:eGFP小鼠的培养脊髓神经元中结合使用松散膜片钳(LP)和全细胞膜片钳(WC)电生理技术,对源自单个GABA/甘氨酸共同释放突触的微小抑制性突触后电流(mIPSC)进行采样,并开发一种建模方法来说明随着甘氨酸在囊泡中取代GABA,mIPSC表型的逐渐变化。如果VIAAT能够摄取这两种氨基酸,则预测会有一致的GABA/甘氨酸平衡,我们测试了来自单个终末的囊泡胞吐作用是否会引发均匀的混合mIPSC群体。我们记录了18个单个突触的mIPSC,并在采样的18个突触中的4个中检测到仅含甘氨酸的mIPSC。其余的(14/18)是共同释放的突触,其中有很大比例的混合GABA/甘氨酸mIPSC具有特征性的双相衰减。大多数(9/14)共同释放的突触没有均匀的表型,而是通过混合和纯mIPSC的组合发出信号,这表明在单个囊泡水平上GABA和甘氨酸的装载和/或储存存在变异性。我们的模型预测,当甘氨酸在突触囊泡中取代GABA时,mIPSC的峰值幅度和电荷转移之间的重新分布有助于维持抑制强度,同时提高信号的时间精度。