Section of Molecular Virology, Imperial College London, London, United Kingdom.
The Roslin Institute, Royal (Dick) School of Veterinary Studies, University of Edinburgh, Midlothian, United Kingdom.
Elife. 2019 Jun 4;8:e45066. doi: 10.7554/eLife.45066.
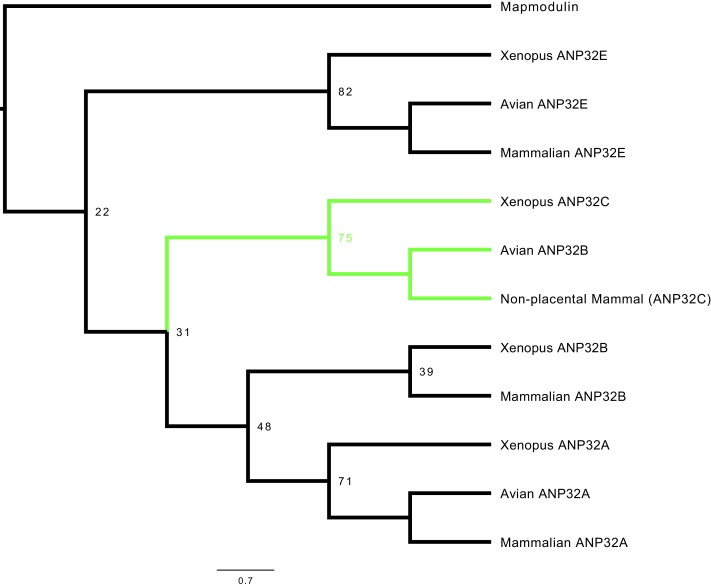
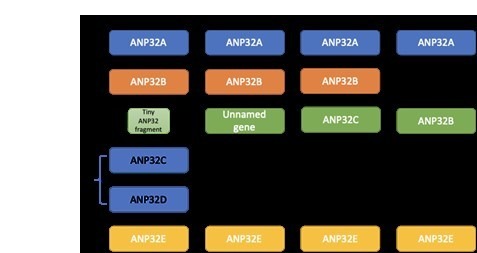
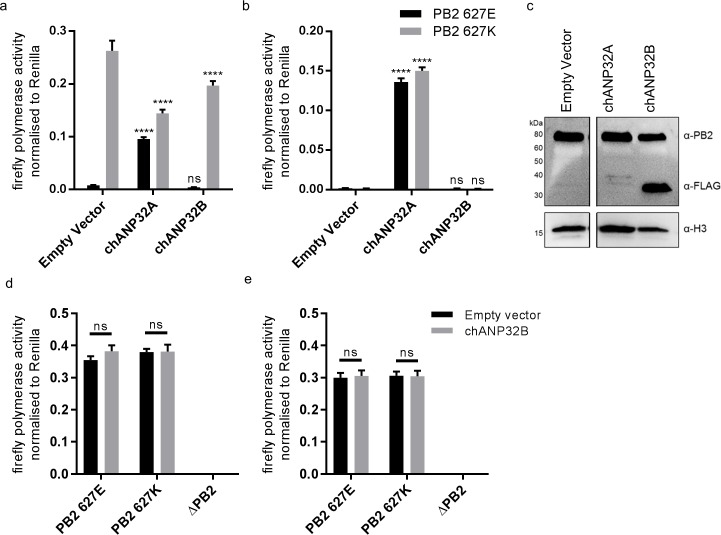
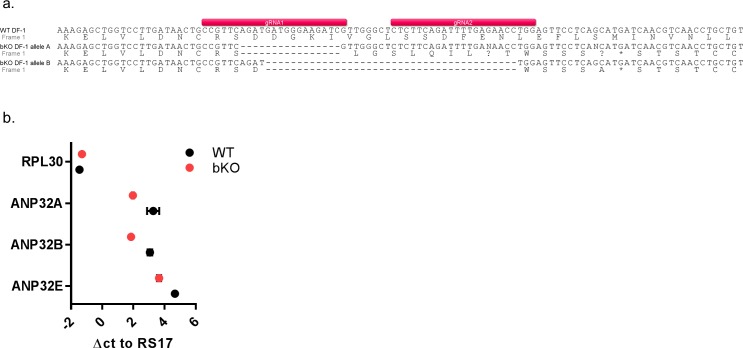
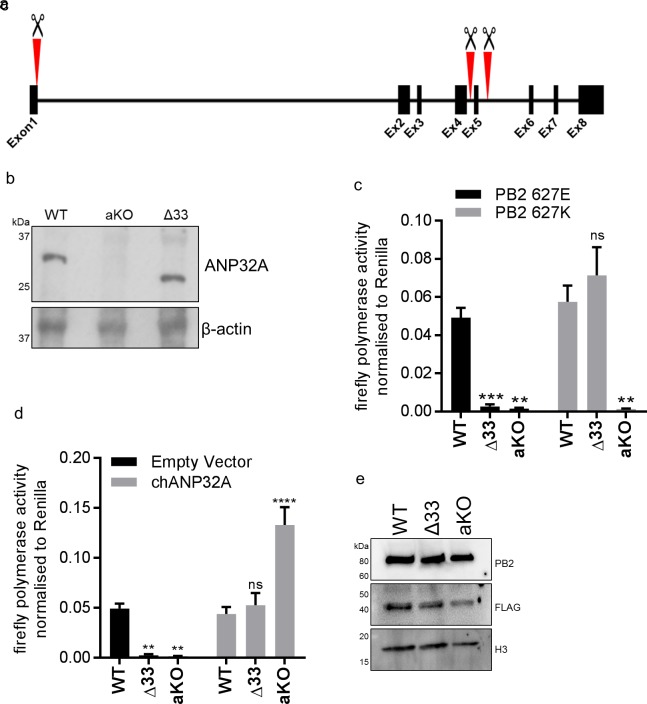
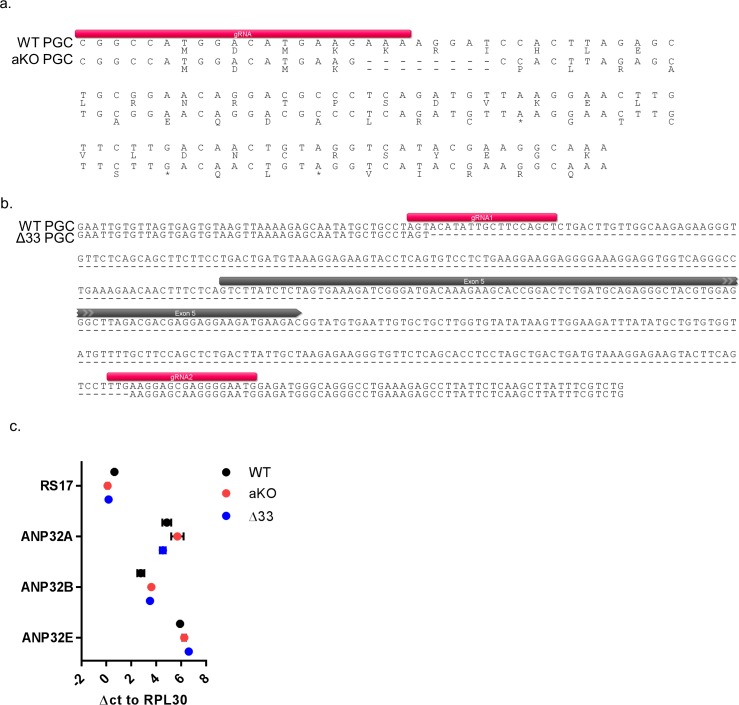
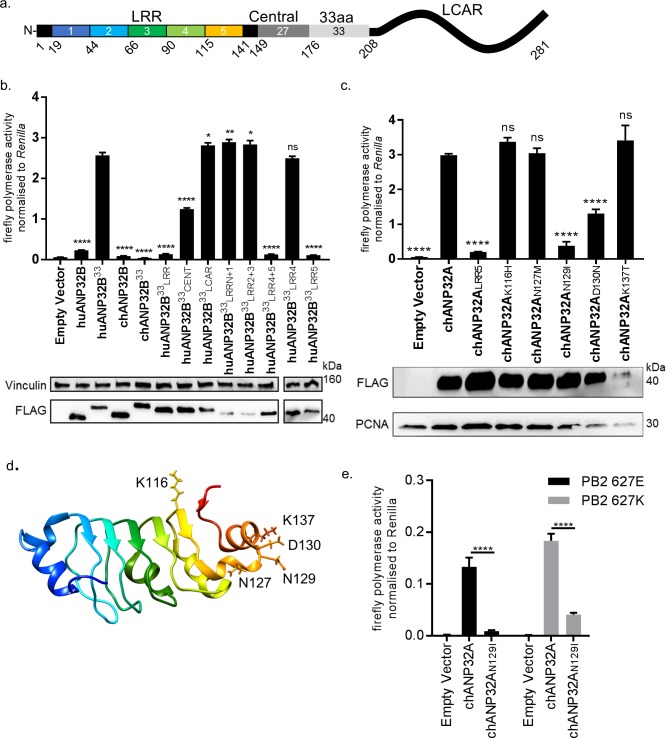
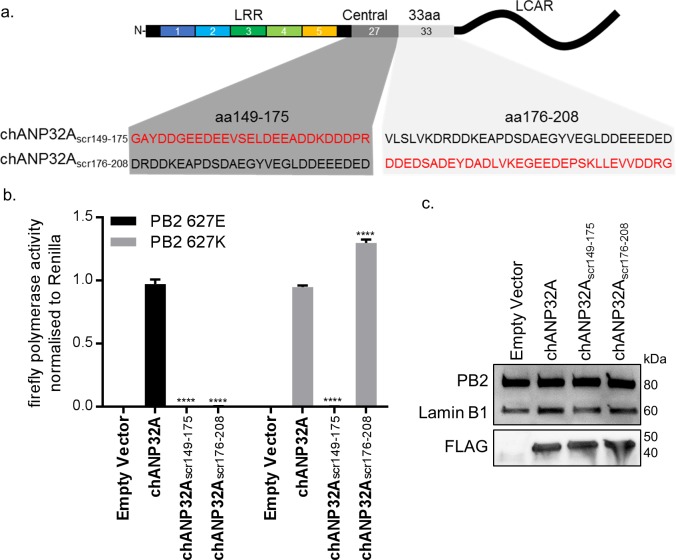
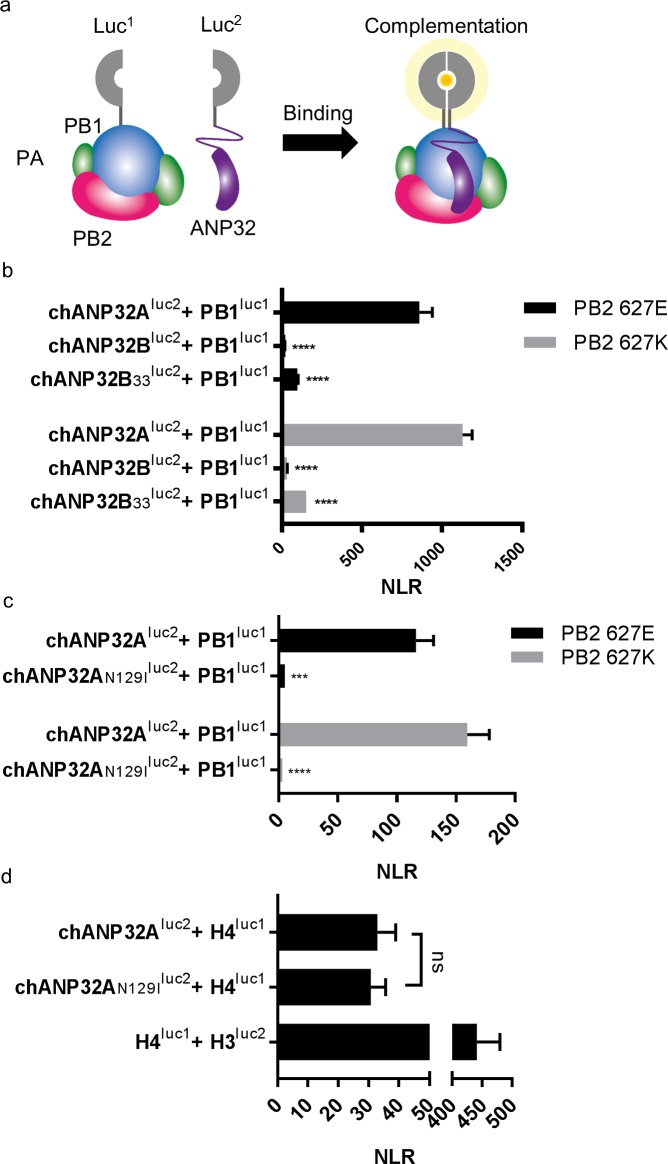
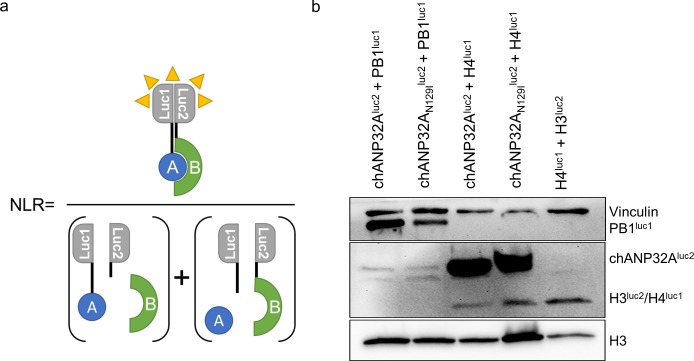
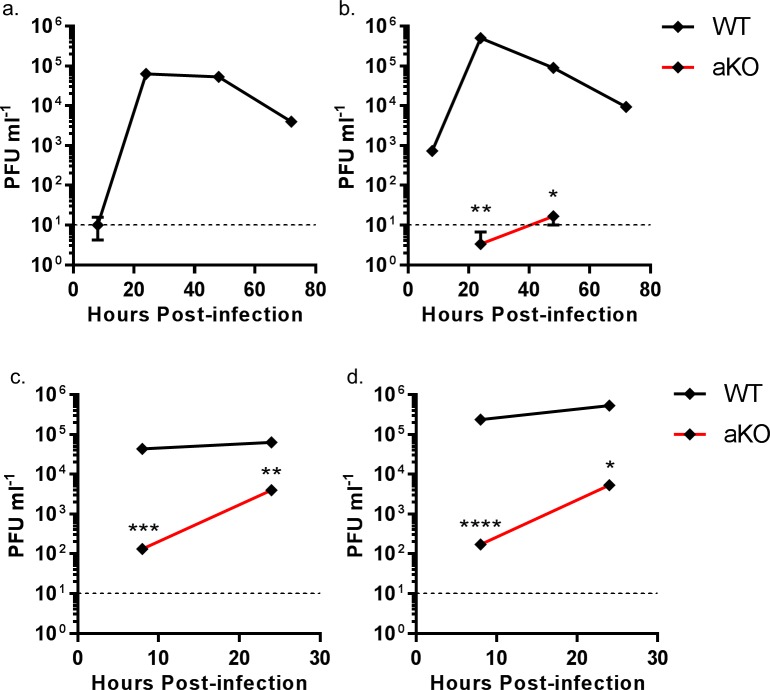
Influenza A viruses (IAV) are subject to species barriers that prevent frequent zoonotic transmission and pandemics. One of these barriers is the poor activity of avian IAV polymerases in human cells. Differences between avian and mammalian ANP32 proteins underlie this host range barrier. Human ANP32A and ANP32B homologues both support function of human-adapted influenza polymerase but do not support efficient activity of avian IAV polymerase which requires avian ANP32A. We show here that the gene currently designated as avian ANP32B is evolutionarily distinct from mammalian ANP32B, and that chicken ANP32B does not support IAV polymerase activity even of human-adapted viruses. Consequently, IAV relies solely on chicken ANP32A to support its replication in chicken cells. Amino acids 129I and 130N, accounted for the inactivity of chicken ANP32B. Transfer of these residues to chicken ANP32A abolished support of IAV polymerase. Understanding ANP32 function will help develop antiviral strategies and aid the design of influenza virus resilient genome edited chickens.
甲型流感病毒(IAV)受到物种屏障的限制,这阻止了它们在动物间频繁传播和引发大流行。其中一个屏障是禽源 IAV 聚合酶在人类细胞中的活性较差。这种宿主范围的障碍是由禽源和哺乳动物的 ANP32 蛋白之间的差异造成的。人类 ANP32A 和 ANP32B 同源物都支持人类适应的流感聚合酶的功能,但不支持需要禽源 ANP32A 的禽源 IAV 聚合酶的有效活性。我们在这里表明,目前被指定为禽源 ANP32B 的基因在进化上与哺乳动物的 ANP32B 不同,鸡的 ANP32B 甚至不支持人类适应的病毒的 IAV 聚合酶活性。因此,IAV 仅依靠鸡的 ANP32A 来支持其在鸡细胞中的复制。氨基酸 129I 和 130N 导致鸡 ANP32B 无活性。将这些残基转移到鸡的 ANP32A 上,会使 IAV 聚合酶失去支持作用。了解 ANP32 的功能将有助于开发抗病毒策略,并有助于设计具有抗流感病毒能力的基因组编辑鸡。