Lai Xuelei, Daher Hussein, Galien Antonin, Hugouvieux Veronique, Zubieta Chloe
Laboratoire de Physiologie Cellulaire et Végétale, CNRS, Univ. Grenoble Alpes, CEA, INRA, IRIG, Grenoble, France.
Comput Struct Biotechnol J. 2019 Jun 14;17:946-953. doi: 10.1016/j.csbj.2019.06.014. eCollection 2019.
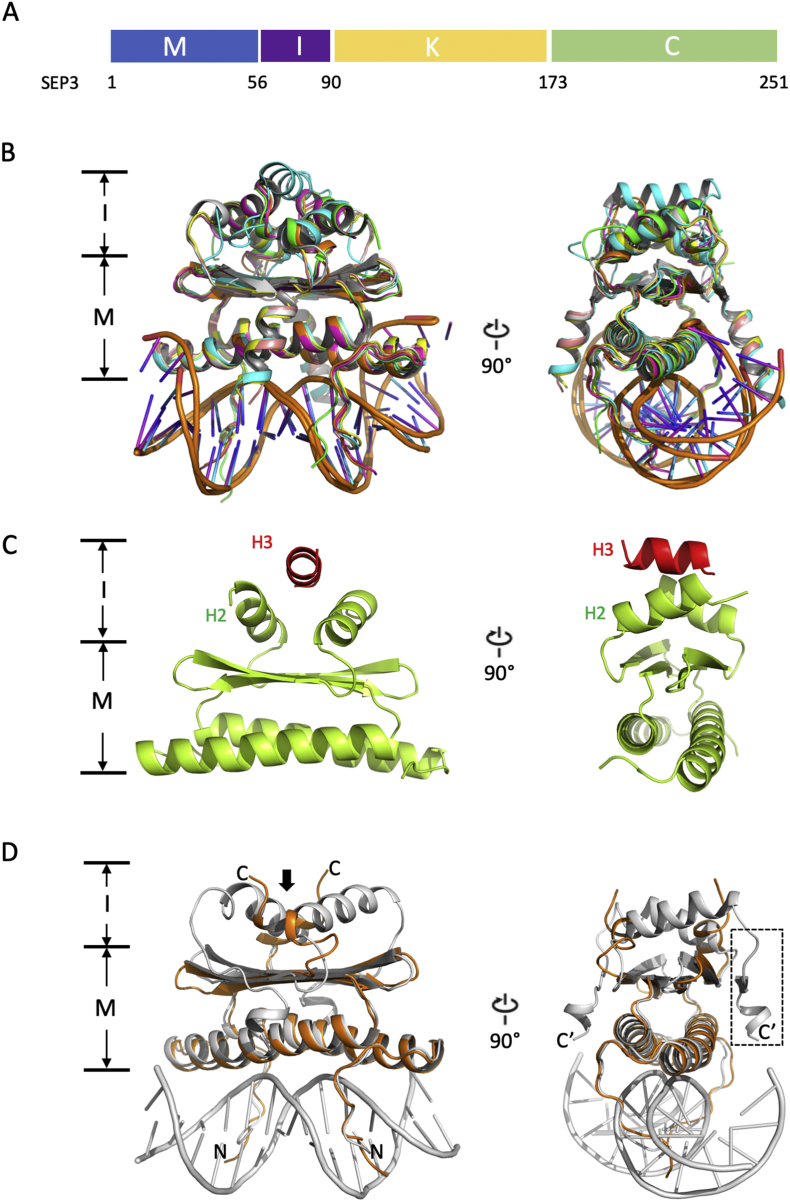
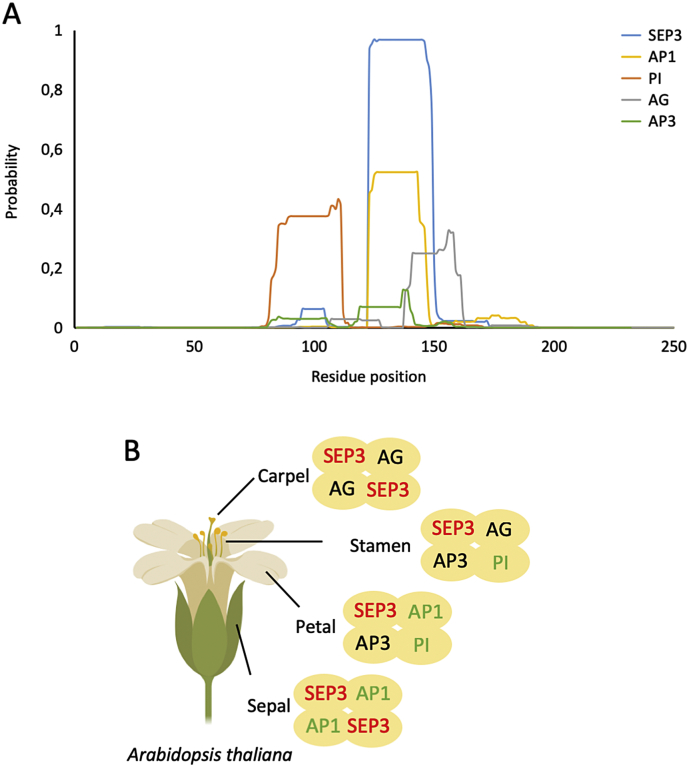
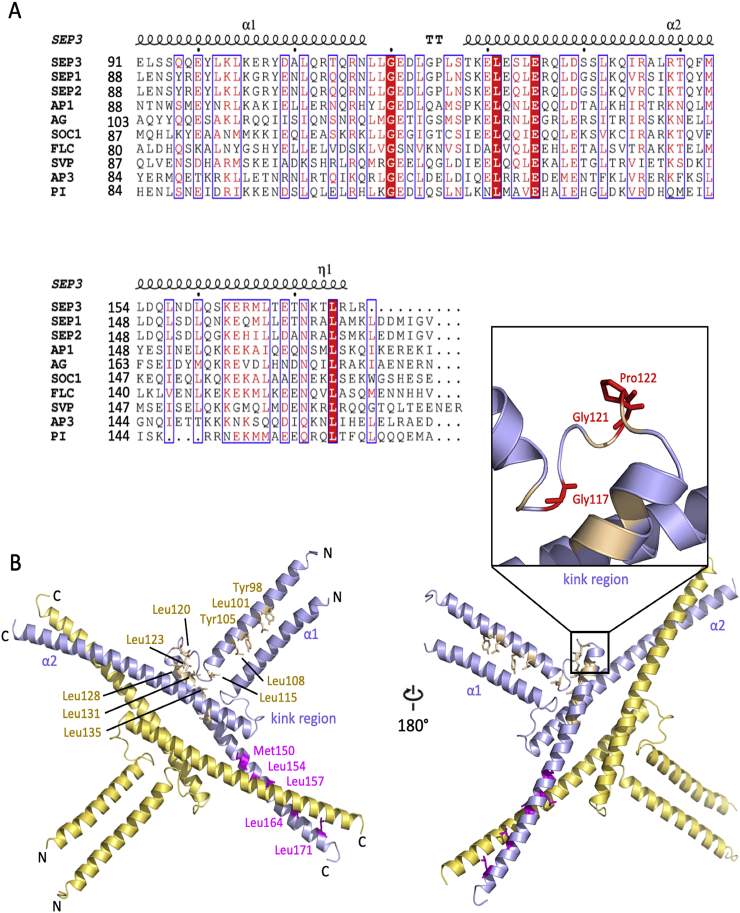
MADS transcription factors (TFs) are DNA binding proteins found in almost all eukaryotes that play essential roles in diverse biological processes. While present in animals and fungi as a small TF family, the family has dramatically expanded in plants over the course of evolution, with the model flowering plant, , possessing over 100 type I and type II MADS TFs. All MADS TFs contain a core and highly conserved DNA binding domain called the MADS or M domain. Plant MADS TFs have diversified this domain with plant-specific auxiliary domains. Plant type I MADS TFs have a highly diverse and largely unstructured Carboxy-terminal (C domain), whereas type II MADS have added oligomerization domains, called Intervening (I domain) and Keratin-like (K domain), in addition to the C domain. In this mini review, we describe the overall structure of the type II "MIKC" type MADS TFs in plants, with a focus on the K domain, a critical oligomerization module. We summarize the determining factors for oligomerization and provide mechanistic insights on how secondary structural elements are required for oligomerization capability and specificity. Using MADS TFs that are involved in flower organ specification as an example, we provide case studies and homology modeling of MADS TFs complex formation. Finally, we highlight outstanding questions in the field.
MADS转录因子(TFs)是几乎在所有真核生物中都能发现的DNA结合蛋白,在多种生物学过程中发挥着重要作用。虽然在动物和真菌中作为一个小的转录因子家族存在,但在植物进化过程中该家族显著扩张,模式开花植物拥有超过100种I型和II型MADS转录因子。所有MADS转录因子都包含一个核心且高度保守的DNA结合结构域,称为MADS或M结构域。植物MADS转录因子通过植物特有的辅助结构域使该结构域多样化。植物I型MADS转录因子具有高度多样且大多无结构的羧基末端(C结构域),而II型MADS除了C结构域之外还增加了寡聚化结构域,称为中间(I结构域)和角蛋白样(K结构域)。在这篇小型综述中,我们描述了植物中II型“MIKC”型MADS转录因子的整体结构,重点关注K结构域,这是一个关键的寡聚化模块。我们总结了寡聚化的决定因素,并提供了关于寡聚化能力和特异性如何需要二级结构元件的机制见解。以参与花器官特化的MADS转录因子为例,我们提供了MADS转录因子复合物形成的案例研究和同源建模。最后,我们强调了该领域中尚未解决的问题。