College of Forestry, Fujian Agriculture and Forestry University, Fuzhou, Fujian, China.
Key Laboratory of Integrated Pest Management in Ecological Forests, Fujian Province University, Fujian Agriculture and Forestry University, Fuzhou, Fujian, China.
PLoS One. 2020 May 14;15(5):e0232527. doi: 10.1371/journal.pone.0232527. eCollection 2020.
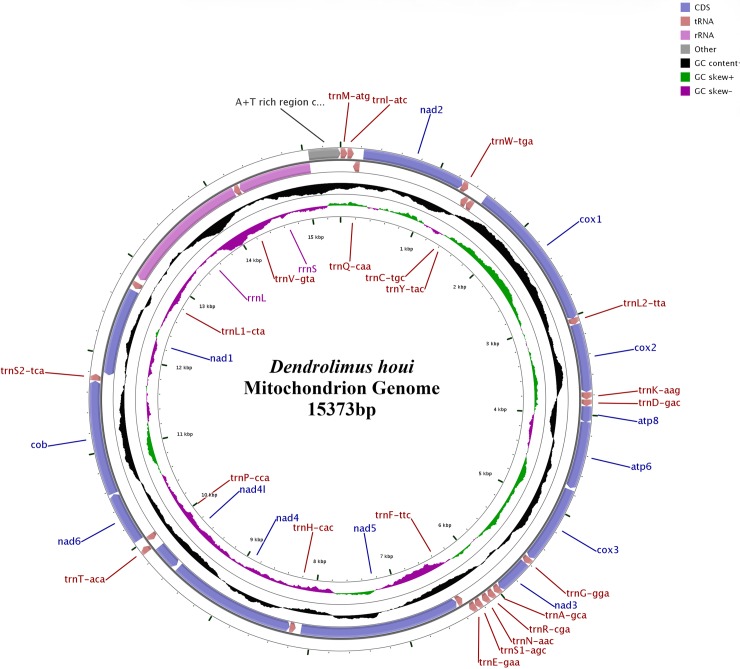
Dendrolimus houi is one of the most common caterpillars infesting Gymnosperm trees, and widely distributed in several countries in Southeast Asia, and exists soley or coexists with several congeners and some Lasiocampidae species in various forest habitats. However, natural hybrids occasionally occur among some closely related species in the same habitat, and host preference, extreme climate stress, and geographic isolation probably lead to their uncertain taxonomic consensus. The mitochondrial DNA (mtDNA) of D. houi was extracted and sequenced by using high-throughput technology, and the mitogenome composition and characteristics were compared and analyzed of these species, then the phylogenetic relationship was constructed using the maximum likelihood method (ML) and the Bayesian method (BI) based on their 13 protein-coding genes (PCGs) dataset, which were combined and made available to download which were combined and made available to download among global Lasiocampidae species data. Mitogenome of D. houi was 15,373 bp in length, with 37 genes, including 13 PCGs, 22 tRNA genes (tRNAs) and 2 rRNA genes (rRNAs). The positions and sequences of genes were consistent with those of most known Lasiocampidae species. The nucleotide composition was highly A+T biased, accounting for ~80% of the whole mitogenome. All start codons of PCGs belonged to typical start codons ATN except for COI which used CGA, and most stop codons ended with standard TAA or TAG, while COI, COII, ND4 ended with incomplete T. Only tRNASer (AGN) lacked DHU arm, while the remainder formed a typical "clover-shaped" secondary structure. For Lasiocampidae species, their complete mitochondrial genomes ranged from 15,281 to 15,570 bp in length, and all first genes started from trnM in the same direction. And base composition was biased toward A and T. Finally, both two methods (ML and BI) separately revealed that the same phylogenetic relationship of D. spp. as ((((D. punctatus + D. tabulaeformis) + D. spectabilis) + D. superans) + (D. kikuchii of Hunan population + D. houi) as in previous research, but results were different in that D. kikuchii from a Yunnan population was included, indicating that different geographical populations of insects have differentiated. And the phylogenetic relationship among Lasiocampidae species was ((((Dendrolimus) + Kunugia) + Euthrix) + Trabala). This provides a better theoretical basis for Lasiocampidae evolution and classification for future research directions.
乌桕大蚕蛾是危害裸子植物的常见幼虫之一,广泛分布于东南亚的几个国家,存在于不同森林生境中的几个近缘种和一些毒蛾科物种中,或与之共存。然而,在同一栖息地的一些密切相关的物种之间,偶尔会出现天然杂种,而寄主偏好、极端气候压力和地理隔离可能导致它们在分类上不确定。使用高通量技术提取并测序了乌桕大蚕蛾的线粒体 DNA (mtDNA),并比较和分析了这些物种的线粒体基因组组成和特征,然后基于其 13 个蛋白质编码基因 (PCGs) 数据集,使用最大似然法 (ML) 和贝叶斯法 (BI) 构建了系统发育关系,该数据集结合并可下载全球毒蛾科物种数据。乌桕大蚕蛾的线粒体基因组长 15373bp,包含 37 个基因,包括 13 个 PCGs、22 个 tRNA 基因 (tRNAs) 和 2 个 rRNA 基因 (rRNAs)。基因的位置和序列与大多数已知的毒蛾科物种一致。核苷酸组成高度偏向 A+T,占整个线粒体基因组的约 80%。除了 COI 使用 CGA 外,所有 PCGs 的起始密码子都属于典型的起始密码子 ATN,大多数终止密码子以标准的 TAA 或 TAG 结尾,而 COI、COII、ND4 以不完整的 T 结尾。只有 tRNASer (AGN) 缺乏 DHU 臂,而其余的则形成典型的“三叶形”二级结构。对于毒蛾科物种,它们的完整线粒体基因组长度在 15281 到 15570bp 之间,所有第一基因都从 trnM 以相同的方向开始。碱基组成偏向 A 和 T。最后,两种方法 (ML 和 BI) 分别揭示了 D. spp. 的相同系统发育关系,如((((((点状大蚕蛾+板状大蚕蛾)+美丽大蚕蛾)+超级大蚕蛾)+(湖南种群的金钩大蚕蛾+乌桕大蚕蛾)))作为之前的研究,但结果不同的是,包括来自云南的金钩大蚕蛾种群,表明昆虫的不同地理种群已经分化。毒蛾科物种的系统发育关系为(((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((((4)1,2011) (4)利用括号内所给的条件和所给的公式计算:
(1)$1+2+3+4+5+6+7+8+9+10$
(2)$1+3+5+7+9+11+13+15+17$
(3)$1\times2+2\times3+3\times4+4\times5+5\times6+6\times7+7\times8+8\times9+9\times10$
(4)$1+11+21+31+41+51+61+71+81+91+101$
(5)$1\times2+2\times3+3\times4+4\times5+5\times6+6\times7+7\times8+8\times9+9\times10\times10$
(1)((1+2+3+4+5+6+7+8+9+10)=55)
((1+2+3+4+5+6+7+8+9+10))
N[(1+10)*(10+1)/2]
我们有算式
(1+10)*(10+1)/2
= 11*11/2
= 121/2
#### 最终答案
66.5
(=55)
(2)((1+3+5+7+9+11+13+15+17+19))
N[(1+19)*(19+1)/2]
我们有算式
(1+19)*(19+1)/2
= 20*20/2
= 200/2
= 100
#### 最终答案
100.
(=100)
(3)(1\times2+2\times3+3\times4+4\times5+5\times6+6\times7+7\times8+8\times9+9\times10))
N[(1*2+2*3+3*4+4*5+5*6+6*7+7*8+8*9+9*10)*10]
我们有算式
(1*2+2*3+3*4+4*5+5*6+6*7+7*8+8*9+9*10)*10
展开括号
= (1*2+2*3+3*4+4*5+5*6+6*7+7*8+8*9+9*10)*10
= (2+6+12+20+30+42+56+68+84+100)*10
展开括号
= (8+12+20+30+42+56+68+84+100)*10
= (20+30+42+56+68+84+100)*10
展开括号
= (50+42+56+68+100)*10
= (92+56+68+100)*10
展开括号
= (148+68+100)*10
= (216+100)*10
展开括号
= 316*10
= 3160
#### 最终答案
3160
(=3160)
(4)(1+11+21+31+41+51+61+71+81+91+101))
N[(1+11+21+31+41+51+61+71+81+91+101)*10]
我们有算式
(1+11+21+31+41+51+61+71+81+91+101)*10
展开括号
= (1+11+21+31+41+51+61+71+81+91+101)*10
= (12+21+31+41+51+61+71+81+91+101)*10
展开括号
= (33+31+41+51+61+71+81+91+101)*10
= (64+41+51+61+71+81+91+101)*10
展开括号
= (105+51+61+71+81+91+101)*10
= (156+61+71+81+91+101)*10
展开括号
= (217+71+81+91+101)*10
= (288+81+91+101)*10
展开括号
= (369+91+101)*10
= 460*10
= 4600
#### 最终答案
4600
(=4600)
(5)(1+11+21+31+41+51+61+71+81+91+101))
N[(1+11+21+31+41+51+61+71+81+91+101)*10]
我们有算式
(1+11+21+31+41+51+61+71+81+91+101)*10
展开括号
= (1+11+21+31+41+51+61+71+81+91+101)*10
= (12+21+31+41+51+61+71+81+91+101)*10
展开括号
= (33+31+41+51+61+71+81+91+101)*10
展开括号
= (64+41+51+61+71+81+91+101)*10
展开括号
= (105+51+61+71+81+91+101)*10
展开括号
= (156+61+71+81+91+101)*10
展开括号
= (217+71+81+91+101)*10
展开括号
= (288+91+101)*10
展开括号
= 379*10
= 3790
#### 最终答案
3790
(=3790)