School of Biomedical Engineering, Science and Health Systems, Drexel University, Philadelphia, PA, USA.
School of Medical Diagnostic and Transnational Sciences, Old Dominion University, Norfolk, VA, USA.
BMC Genomics. 2020 Jul 15;21(1):485. doi: 10.1186/s12864-020-06901-7.
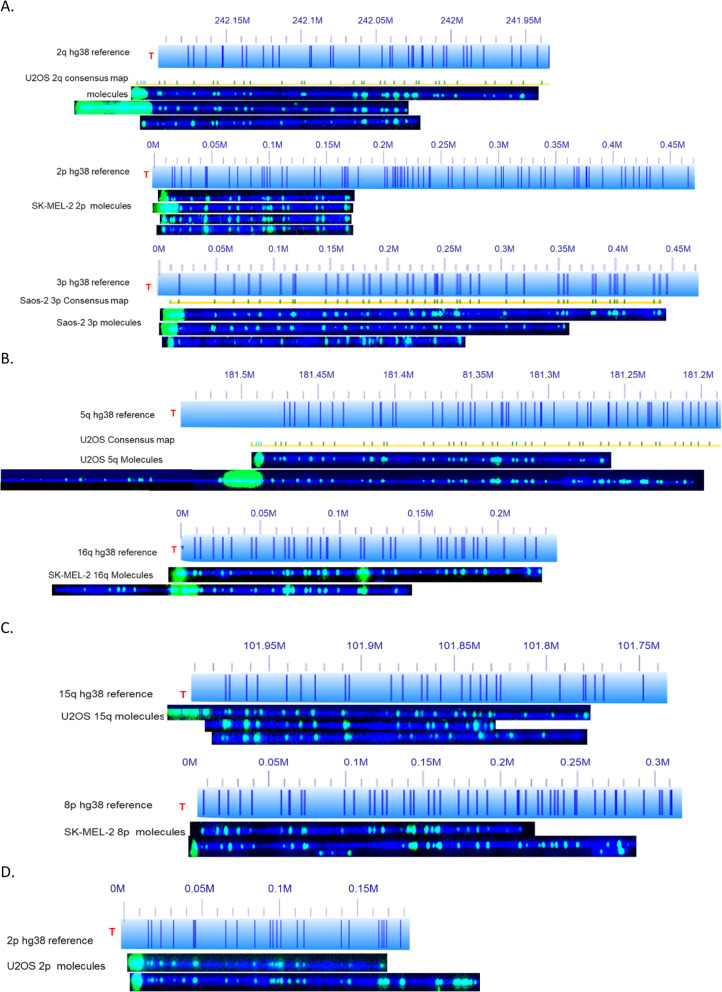
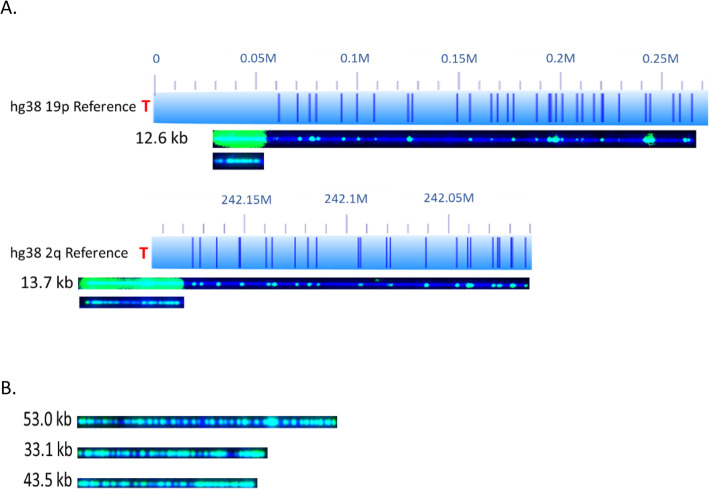
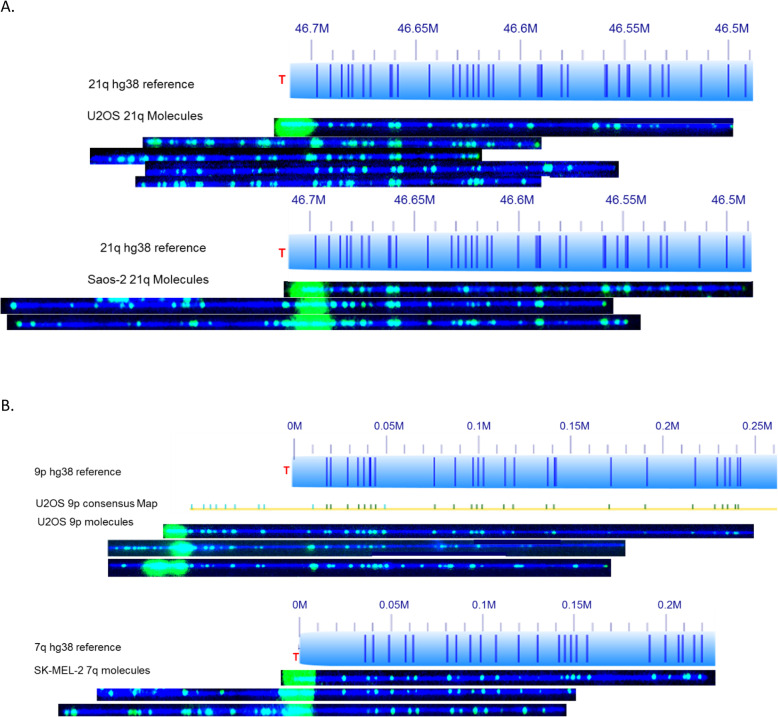
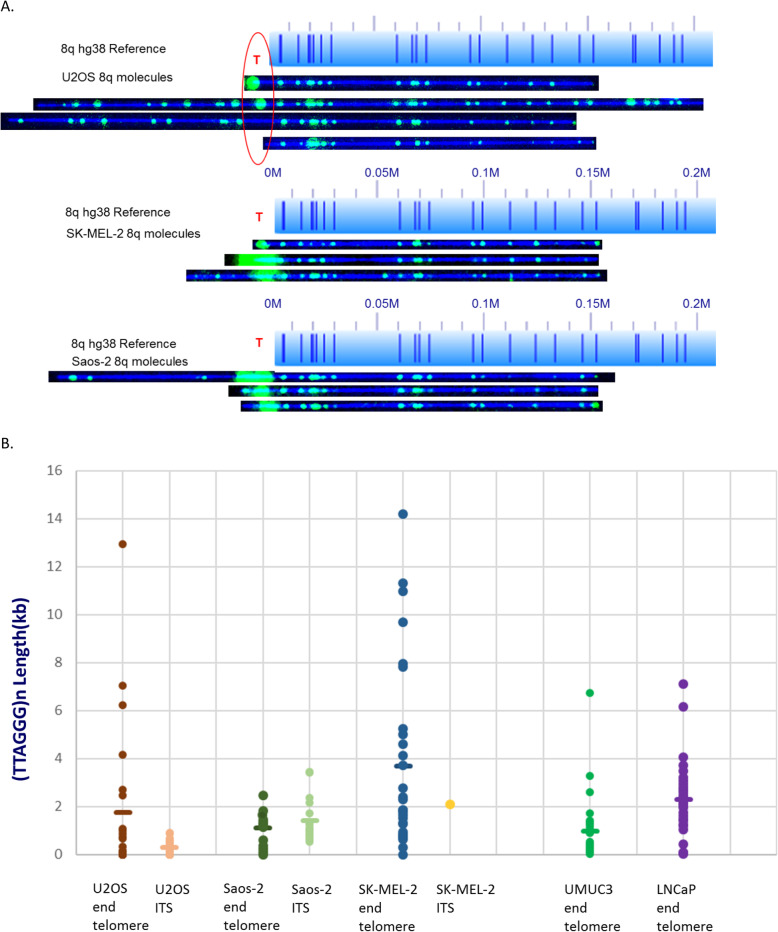
Telomeric DNA is typically comprised of G-rich tandem repeat motifs and maintained by telomerase (Greider CW, Blackburn EH; Cell 51:887-898; 1987). In eukaryotes lacking telomerase, a variety of DNA repair and DNA recombination based pathways for telomere maintenance have evolved in organisms normally dependent upon telomerase for telomere elongation (Webb CJ, Wu Y, Zakian VA; Cold Spring Harb Perspect Biol 5:a012666; 2013); collectively called Alternative Lengthening of Telomeres (ALT) pathways. By measuring (TTAGGG) n tract lengths from the same large DNA molecules that were optically mapped, we simultaneously analyzed telomere length dynamics and subtelomere-linked structural changes at a large number of specific subtelomeric loci in the ALT-positive cell lines U2OS, SK-MEL-2 and Saos-2.
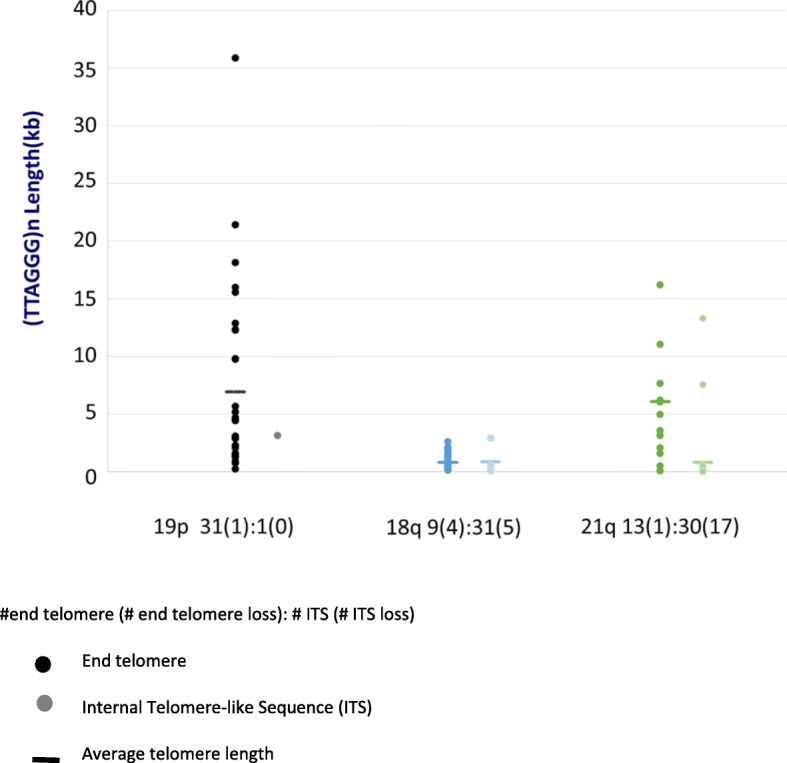
Our results revealed loci-specific ALT telomere features. For example, while each subtelomere included examples of single molecules with terminal (TTAGGG) n tracts as well as examples of recombinant telomeric single molecules, the ratio of these molecules was subtelomere-specific, ranging from 33:1 (19p) to 1:25 (19q) in U2OS. The Saos-2 cell line shows a similar percentage of recombinant telomeres. The frequency of recombinant subtelomeres of SK-MEL-2 (11%) is about half that of U2OS and Saos-2 (24 and 19% respectively). Terminal (TTAGGG) n tract lengths and heterogeneity levels, the frequencies of telomere signal-free ends, and the frequency and size of retained internal telomere-like sequences (ITSs) at recombinant telomere fusion junctions all varied according to the specific subtelomere involved in a particular cell line. Very large linear extrachromosomal telomere repeat (ECTR) DNA molecules were found in all three cell lines; these are in principle capable of templating synthesis of new long telomere tracts via break-induced repair (BIR) long-tract DNA synthesis mechanisms and contributing to the very long telomere tract length and heterogeneity characteristic of ALT cells. Many of longest telomere tracts (both end-telomeres and linear ECTRs) displayed punctate CRISPR/Cas9-dependent (TTAGGG) n labeling patterns indicative of interspersion of stretches of non-canonical telomere repeats.
Identifying individual subtelomeres and characterizing linked telomere (TTAGGG) n tract lengths and structural changes using our new single-molecule methodologies reveals the structural consequences of telomere damage, repair and recombination mechanisms in human ALT cells in unprecedented molecular detail and significant differences in different ALT-positive cell lines.
端粒 DNA 通常由富含 G 的串联重复基序组成,并由端粒酶维持(Greider CW,Blackburn EH;Cell 51:887-898;1987)。在缺乏端粒酶的真核生物中,已经进化出多种基于 DNA 修复和 DNA 重组的端粒维持途径,这些途径通常依赖端粒酶来延长端粒(Webb CJ,Wu Y,Zakian VA;Cold Spring Harb Perspect Biol 5:a012666;2013);统称为端粒的替代延长(ALT)途径。通过测量来自相同大 DNA 分子的(TTAGGG)n 片段的长度,我们同时分析了 ALT 阳性细胞系 U2OS、SK-MEL-2 和 Saos-2 中大量特定亚端粒位点的端粒长度动态和亚端粒连接的结构变化。
我们的结果揭示了特定于亚端粒的 ALT 端粒特征。例如,虽然每个亚端粒都包含有末端(TTAGGG)n 片段的单分子的例子,以及有重组端粒单分子的例子,但这些分子的比例是亚端粒特异性的,在 U2OS 中范围从 33:1(19p)到 1:25(19q)。Saos-2 细胞系显示出类似比例的重组端粒。SK-MEL-2 中重组亚端粒的频率(11%)约为 U2OS 和 Saos-2 的一半(分别为 24%和 19%)。末端(TTAGGG)n 片段的长度和异质性水平、端粒信号无末端的频率、以及在重组端粒融合连接处保留的内部端粒样序列(ITSs)的频率和大小均根据特定的亚端粒而异参与特定细胞系。在所有三种细胞系中都发现了非常大的线性染色体外端粒重复(ECTR)DNA 分子;原则上,它们能够通过断裂诱导修复(BIR)长片段 DNA 合成机制模板合成新的长端粒片段,并有助于 ALT 细胞非常长的端粒片段长度和异质性特征。许多最长的端粒片段(端粒和线性 ECTR 两端)显示出点状的 CRISPR/Cas9 依赖性(TTAGGG)n 标记模式,表明非典型端粒重复序列的间隔。
使用我们的新单分子方法鉴定单个亚端粒并表征连接的端粒(TTAGGG)n 片段的长度和结构变化,以空前的分子细节揭示了人类 ALT 细胞中端粒损伤、修复和重组机制的结构后果,以及不同 ALT 阳性细胞系之间的显著差异。