Cook David E, Kramer H Martin, Torres David E, Seidl Michael F, Thomma Bart P H J
Department of Plant Pathology, Kansas State University, Manhattan, United States.
Laboratory of Phytopathology, Wageningen University & Research, Wageningen, Netherlands.
Elife. 2020 Dec 18;9:e62208. doi: 10.7554/eLife.62208.
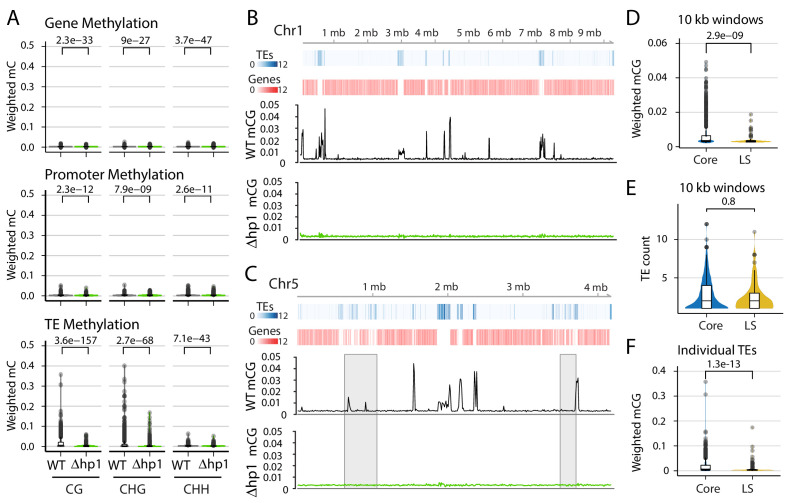
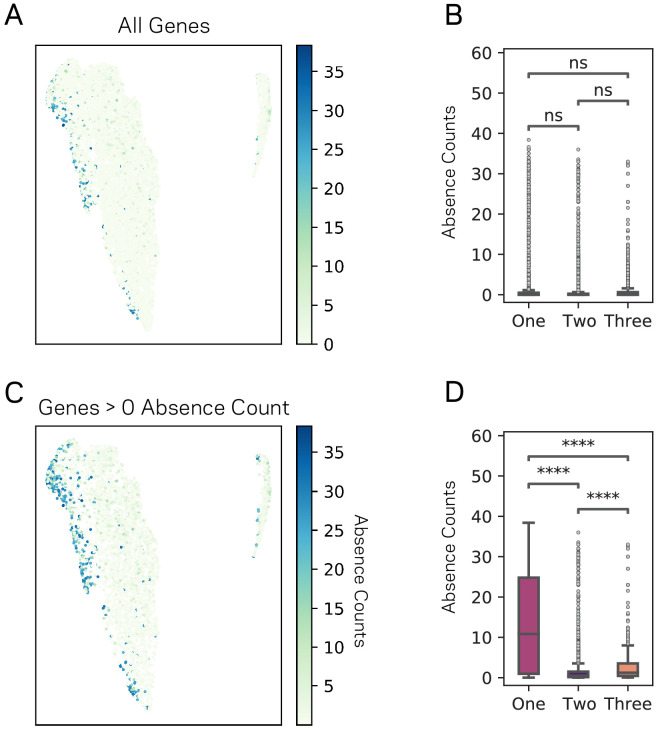
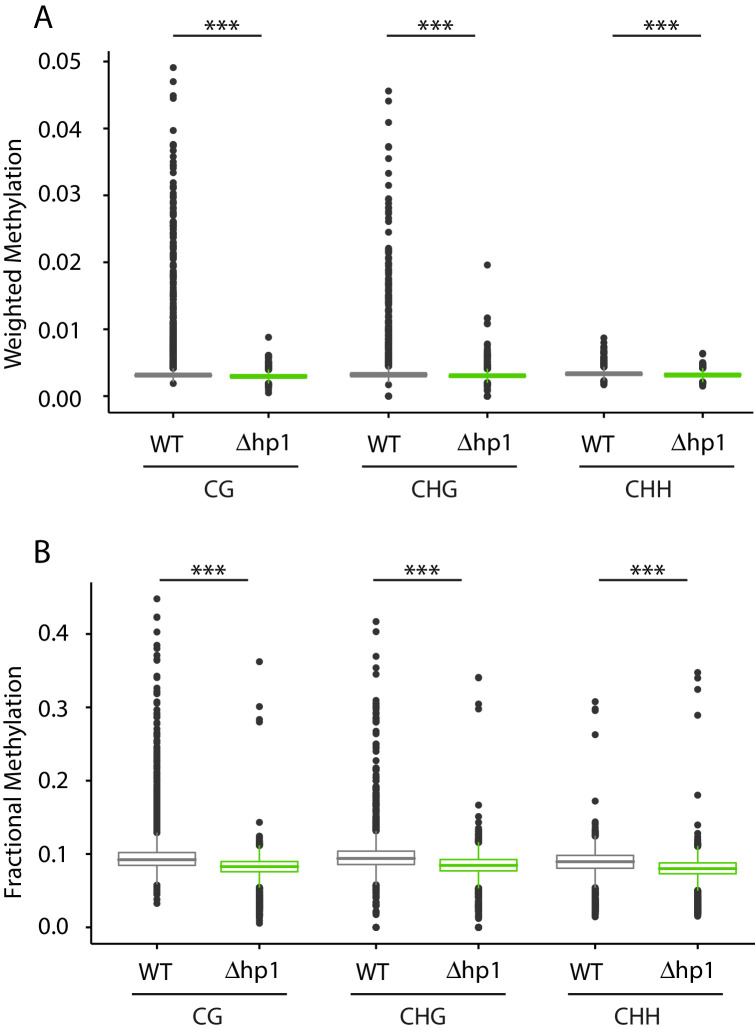
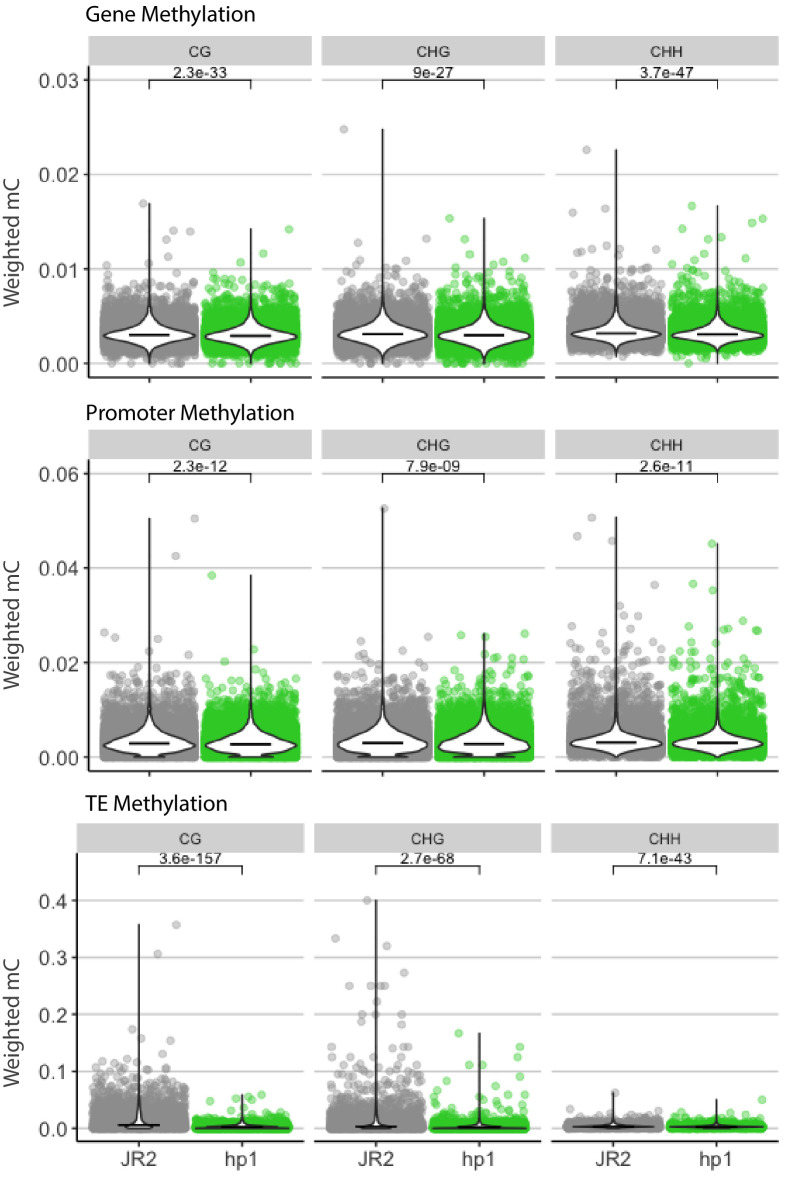
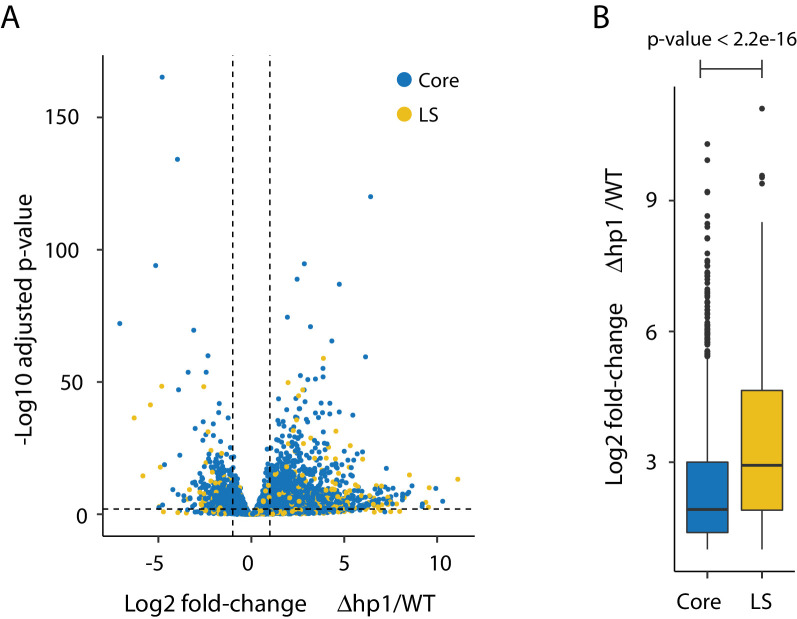
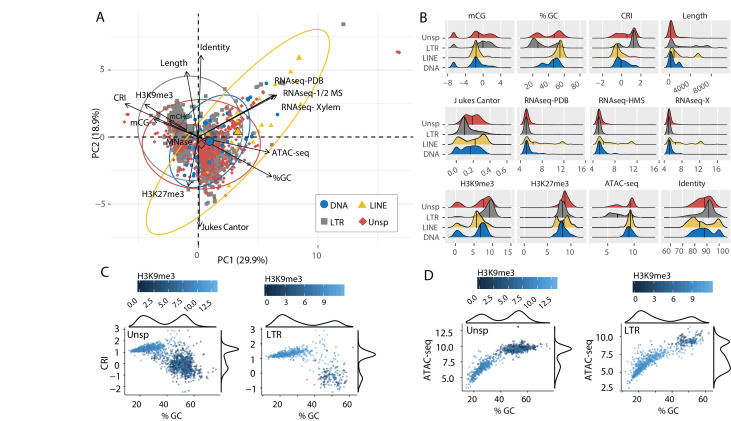
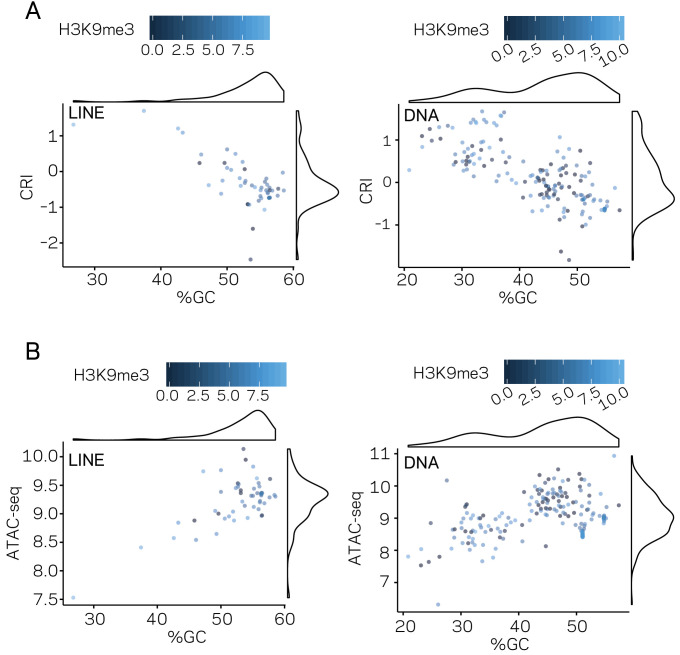
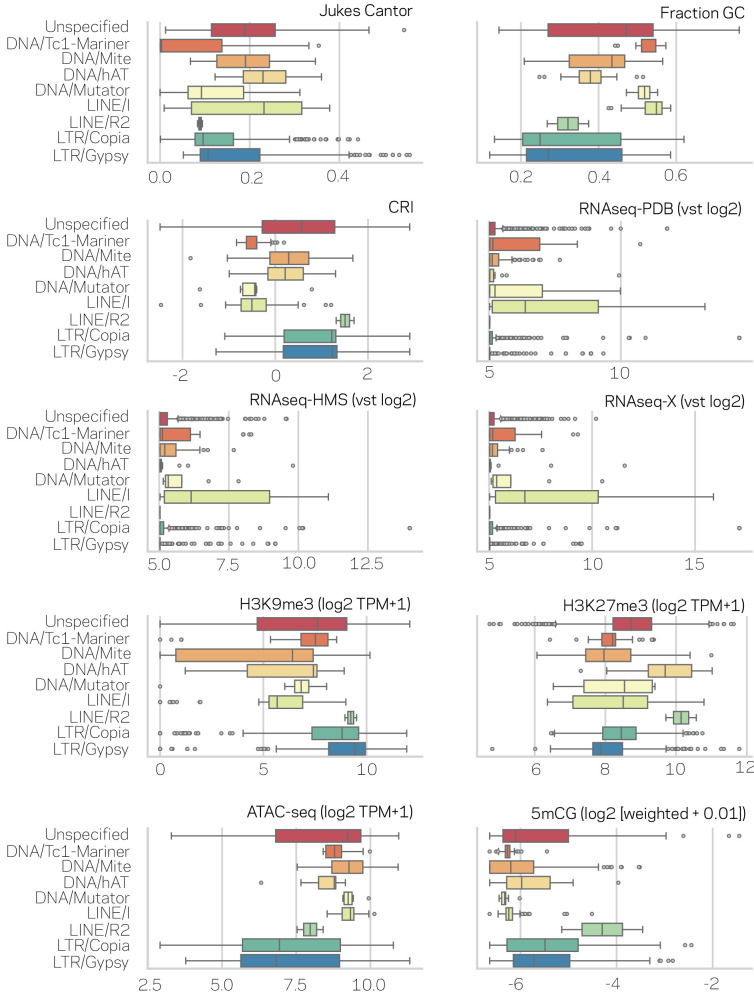
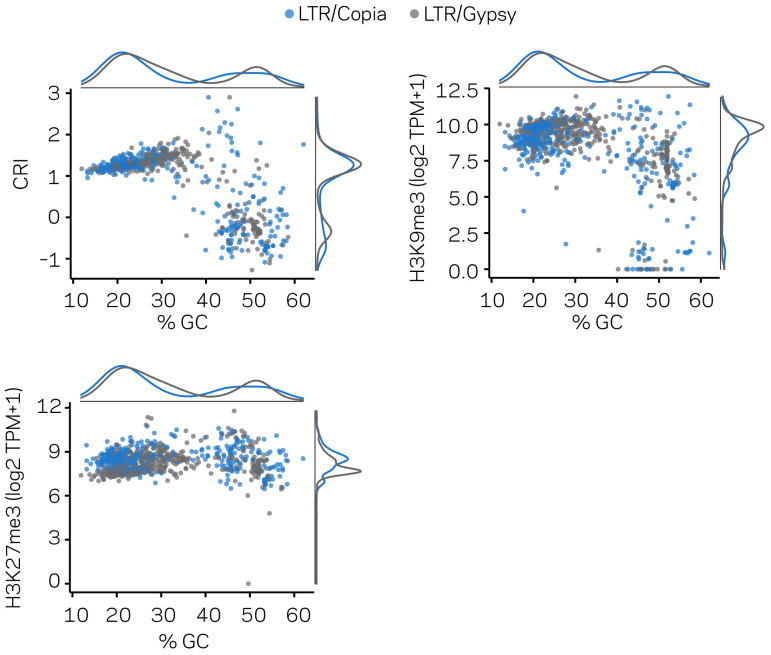
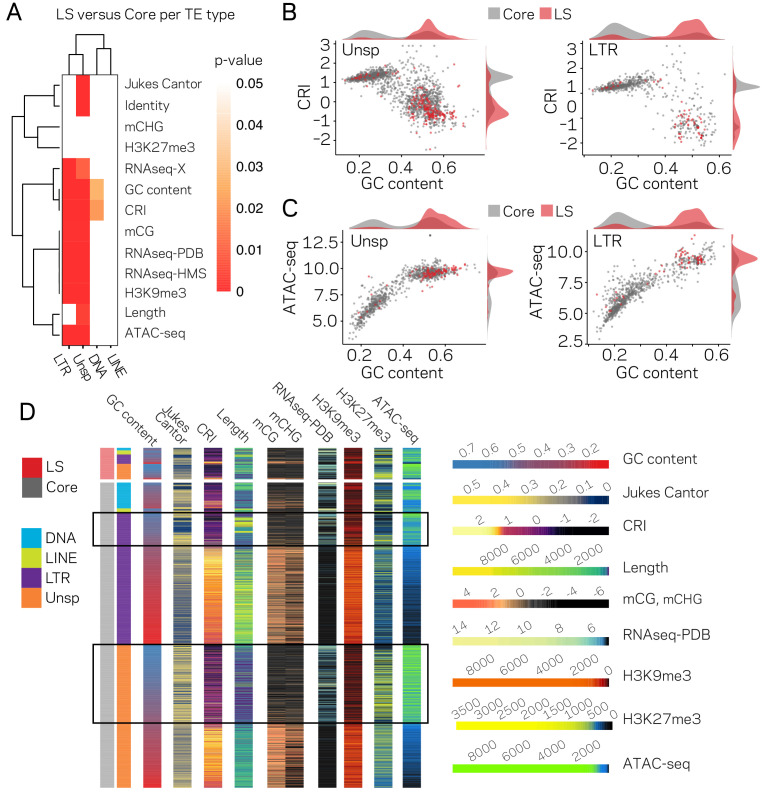
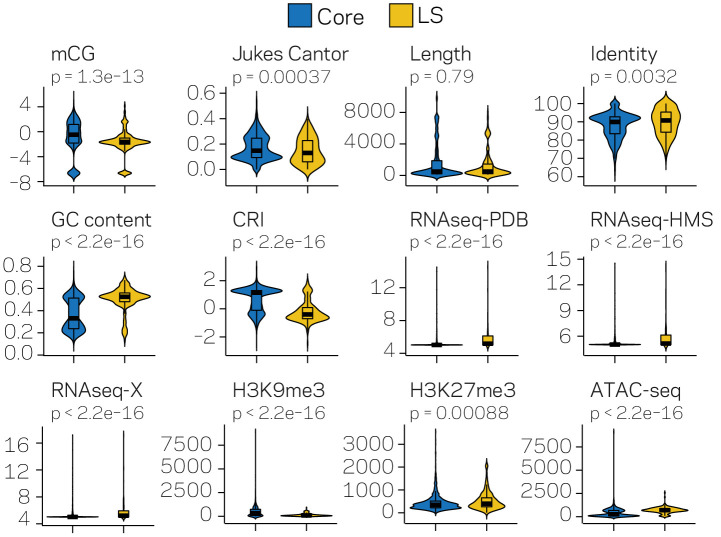
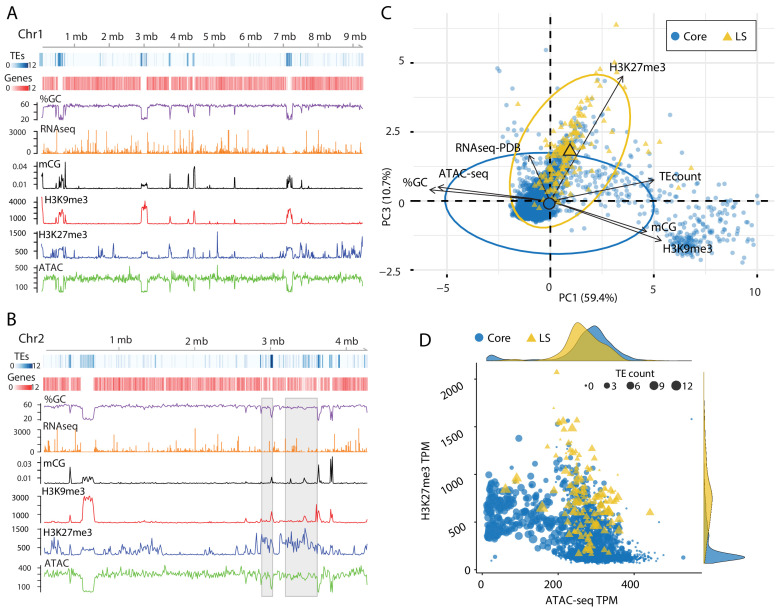
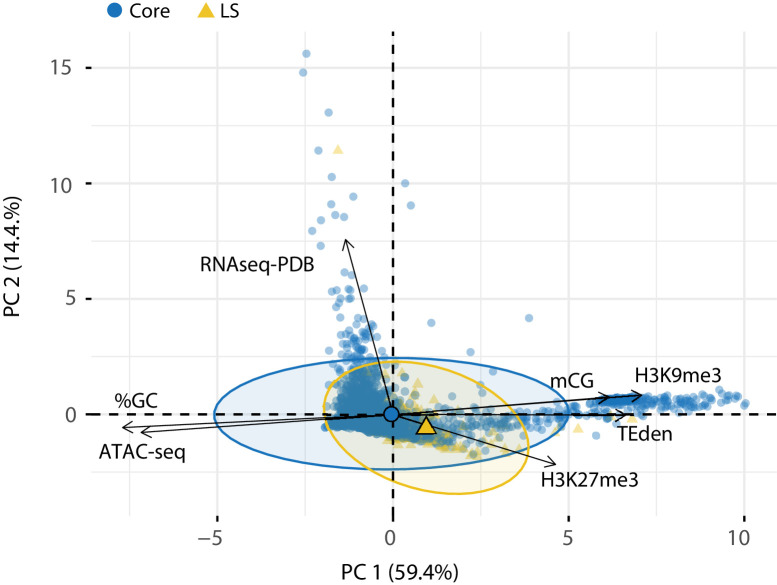
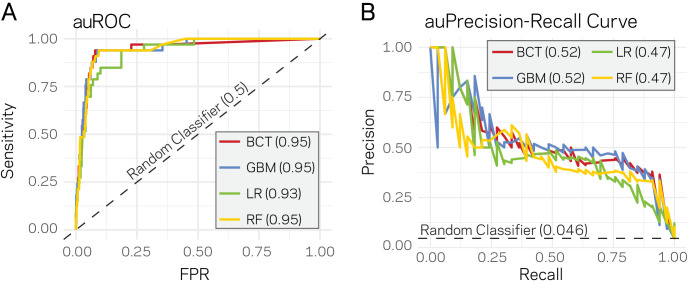
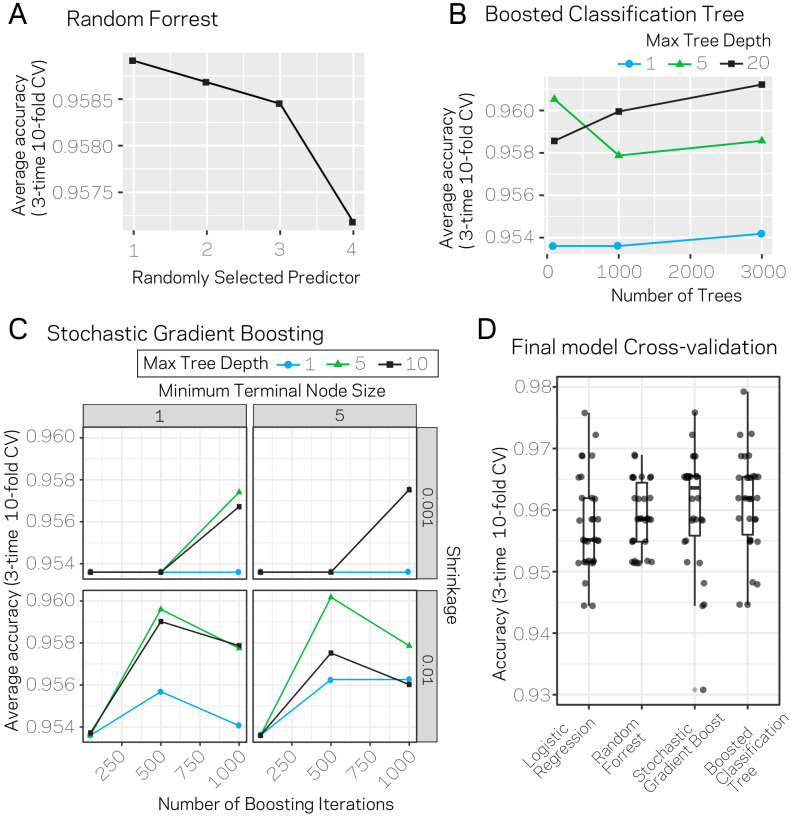
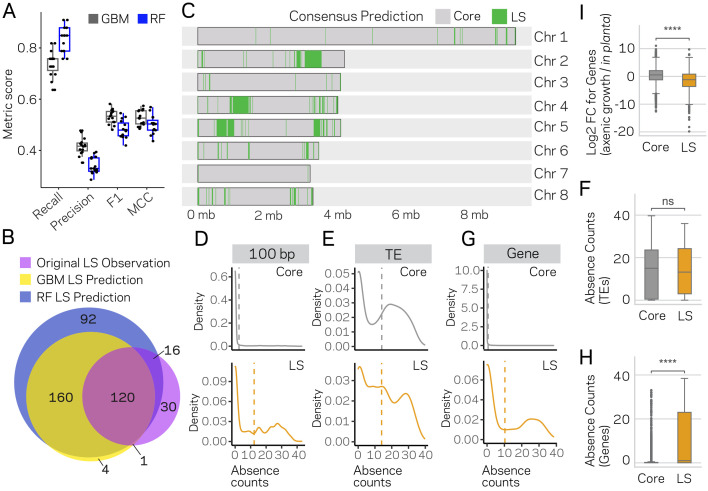
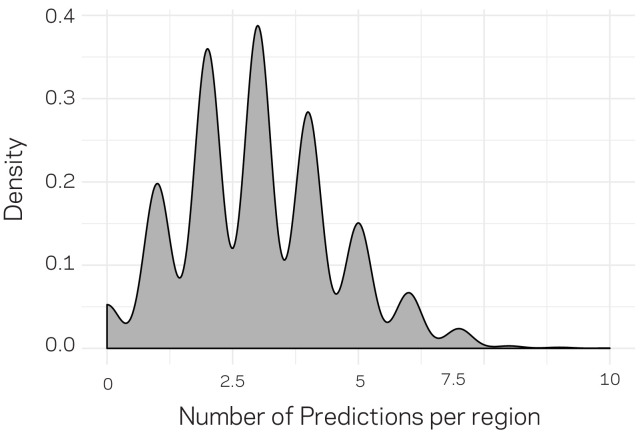
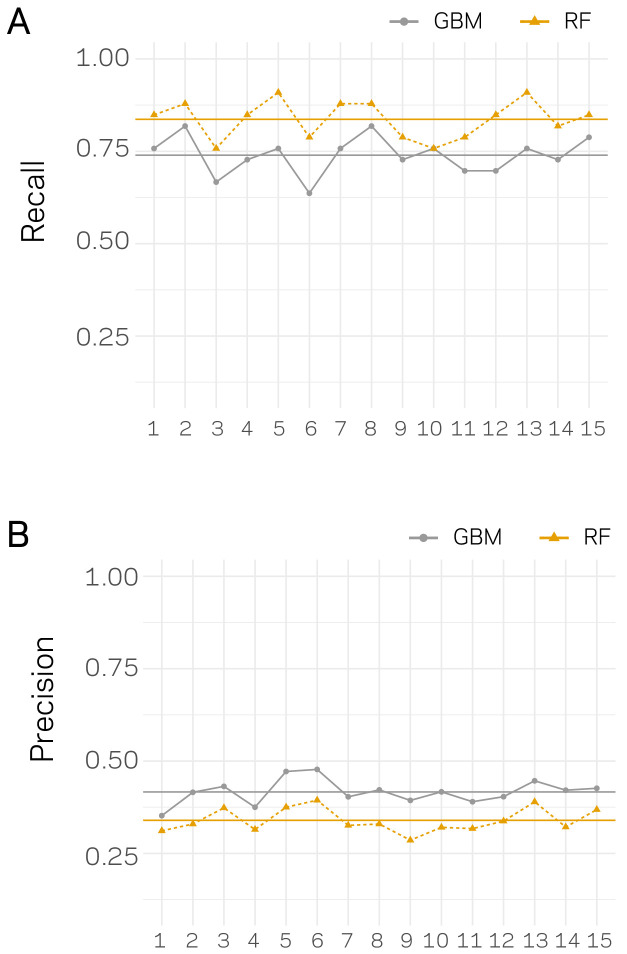
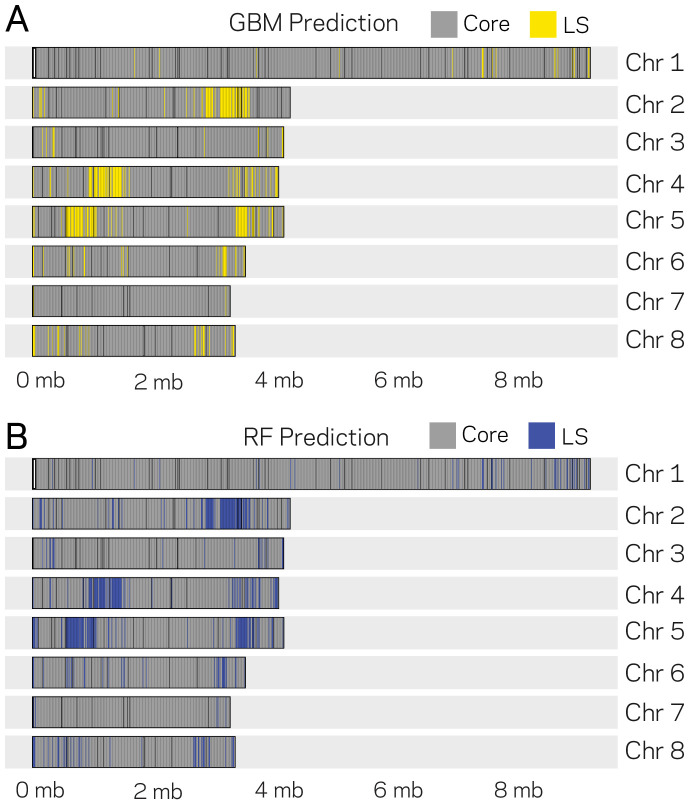
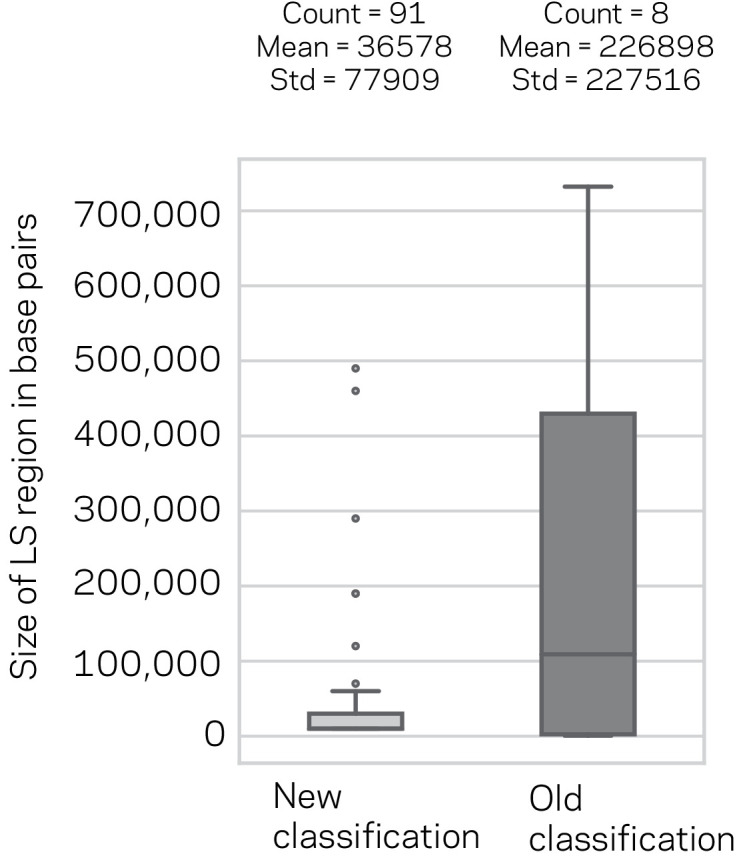
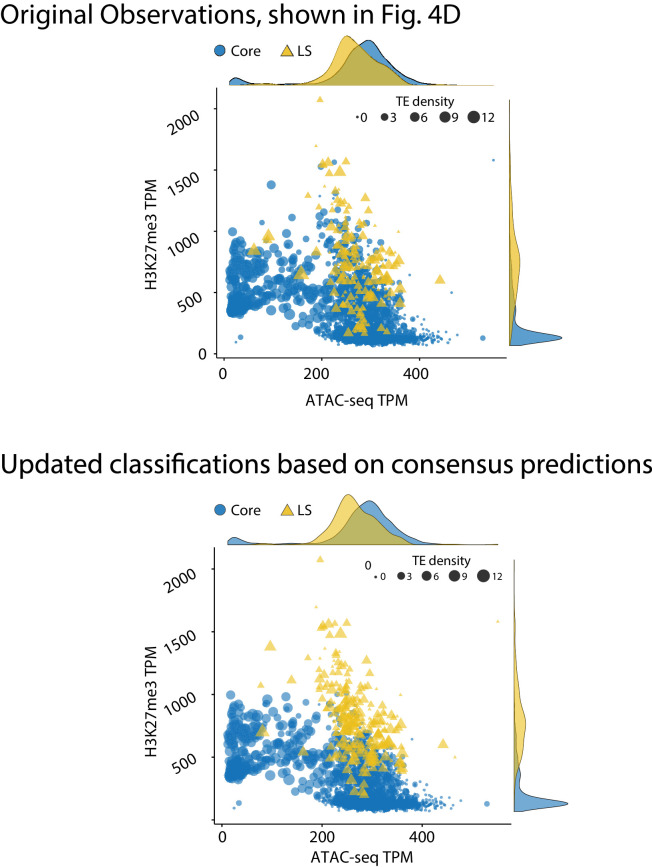
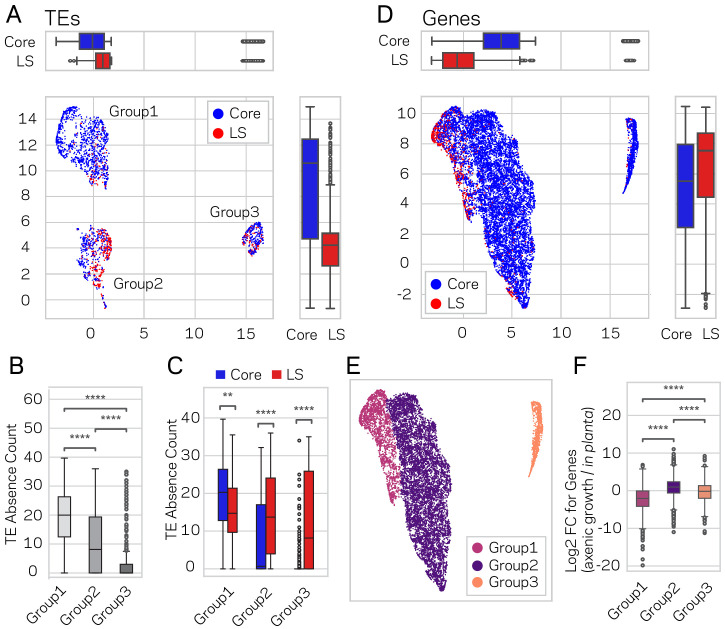
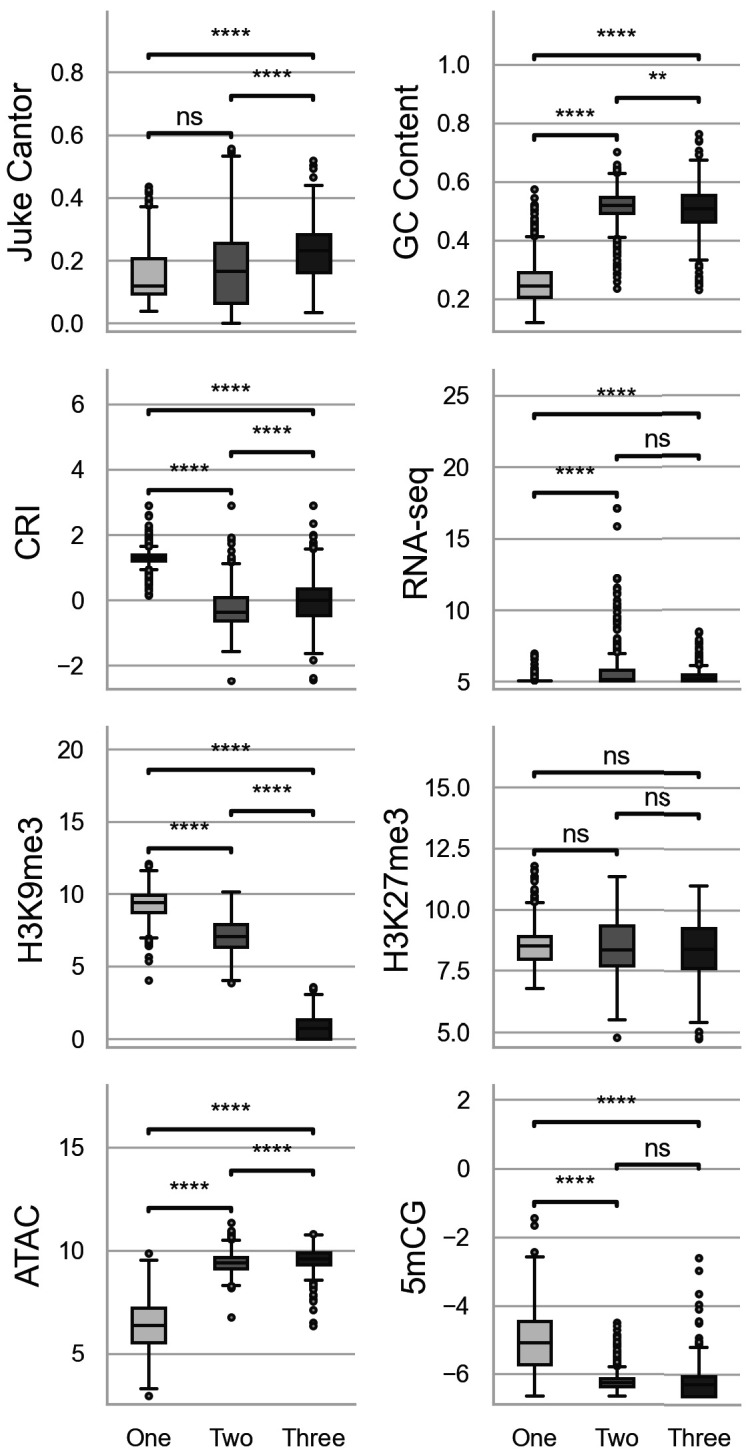
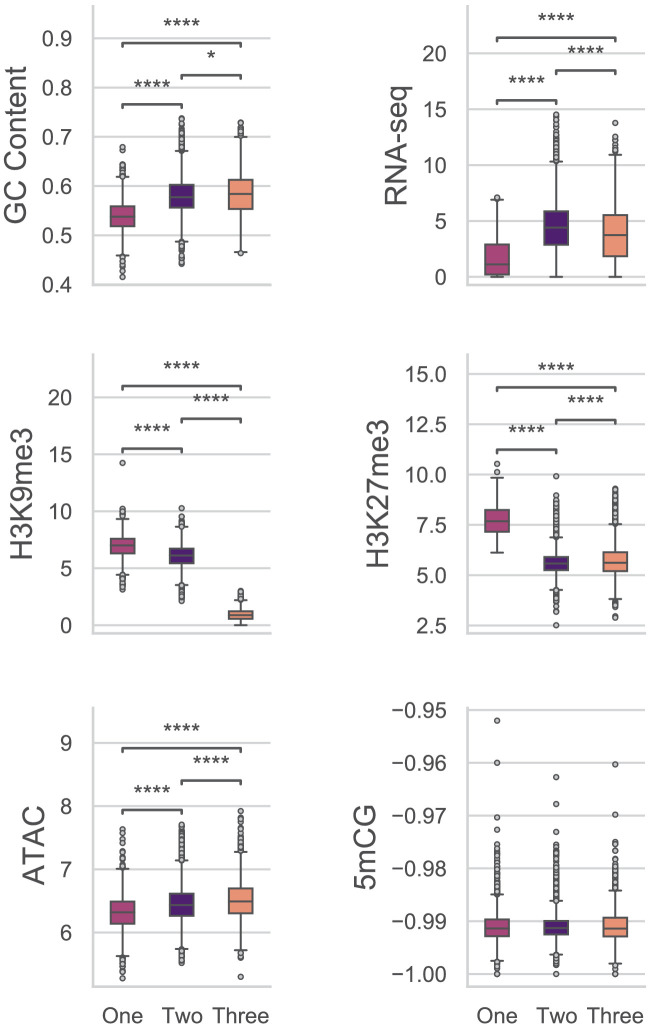
Genomes store information at scales beyond the linear nucleotide sequence, which impacts genome function at the level of an individual, while influences on populations and long-term genome function remains unclear. Here, we addressed how physical and chemical DNA characteristics influence genome evolution in the plant pathogenic fungus . We identified incomplete DNA methylation of repetitive elements, associated with specific genomic compartments originally defined as Lineage-Specific (LS) regions that contain genes involved in host adaptation. Further chromatin characterization revealed associations with features such as H3 Lys-27 methylated histones (H3K27me3) and accessible DNA. Machine learning trained on chromatin data identified twice as much LS DNA as previously recognized, which was validated through orthogonal analysis, and we propose to refer to this DNA as adaptive genomic regions. Our results provide evidence that specific chromatin profiles define adaptive genomic regions, and highlight how different epigenetic factors contribute to the organization of these regions.
基因组储存信息的规模超出了线性核苷酸序列,这在个体层面影响基因组功能,而对种群和长期基因组功能的影响仍不清楚。在这里,我们探讨了物理和化学DNA特征如何影响植物病原真菌的基因组进化。我们发现重复元件的DNA甲基化不完全,这与最初定义为谱系特异性(LS)区域的特定基因组区域相关,这些区域包含参与宿主适应的基因。进一步的染色质特征分析揭示了与诸如H3赖氨酸-27甲基化组蛋白(H3K27me3)和可及DNA等特征的关联。基于染色质数据训练的机器学习识别出的LS DNA是之前所认识到的两倍,这通过正交分析得到了验证,并且我们建议将这种DNA称为适应性基因组区域。我们的结果提供了证据,表明特定的染色质图谱定义了适应性基因组区域,并突出了不同的表观遗传因素如何促成这些区域的组织。