Department of Biotechnology, Pondicherry University, Puducherry, India.
Department of Biology, Indian Institute of Science Education and Research (IISER) Tirupati, Tirupati, India.
Life Sci Alliance. 2020 Dec 29;4(3). doi: 10.26508/lsa.202000699. Print 2021 Mar.
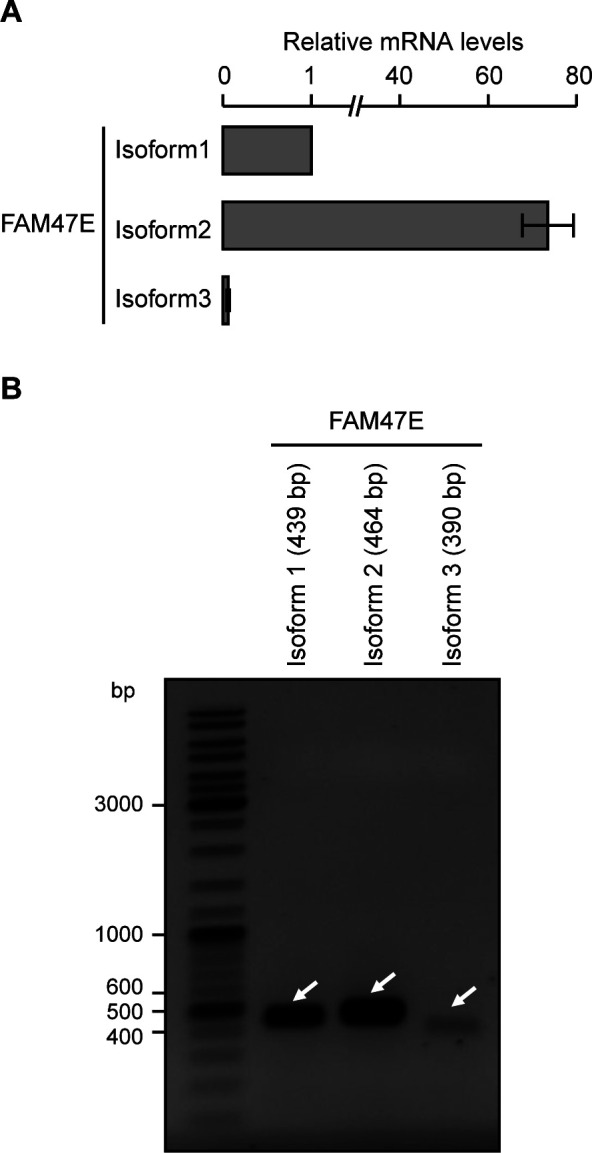
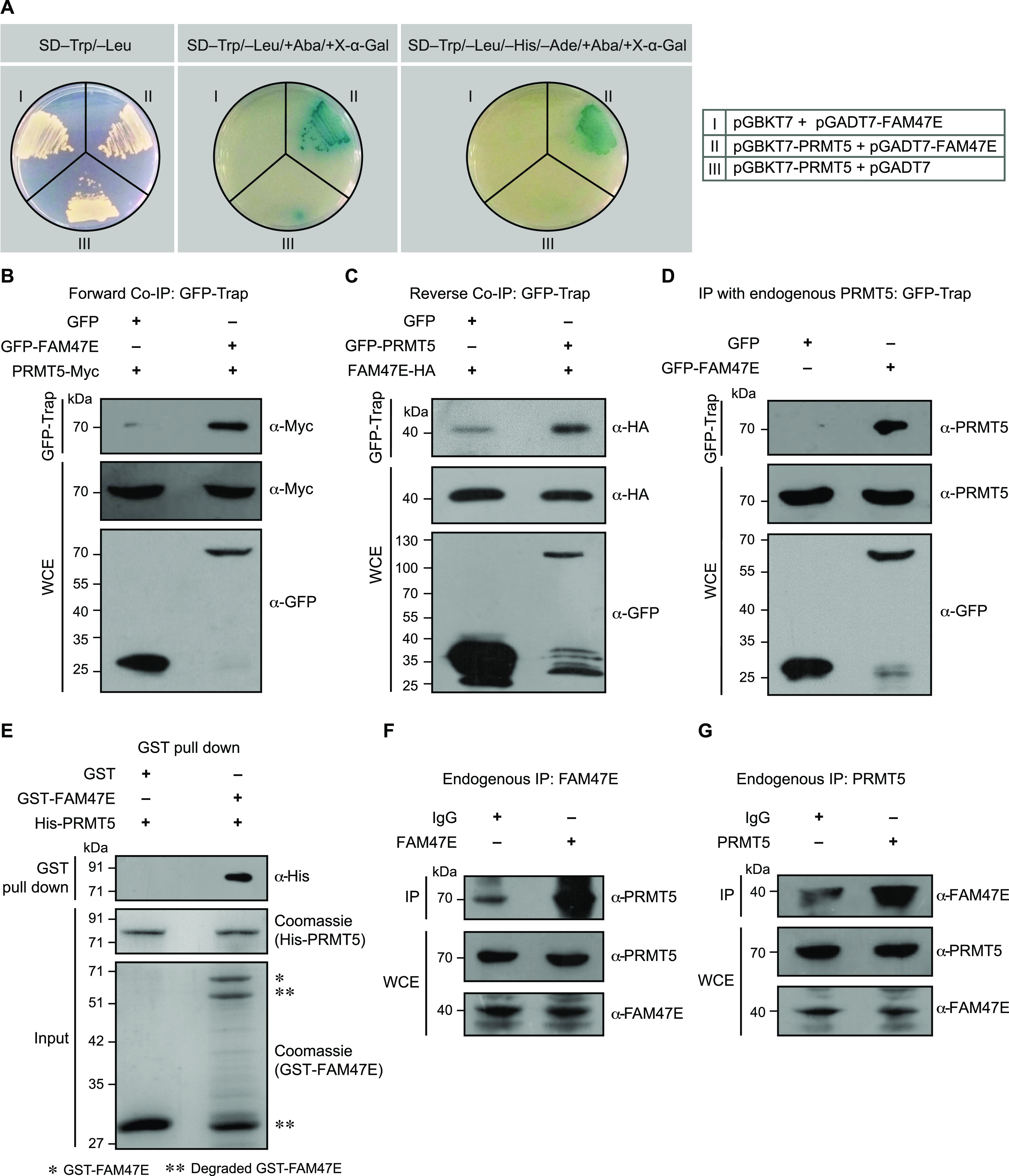
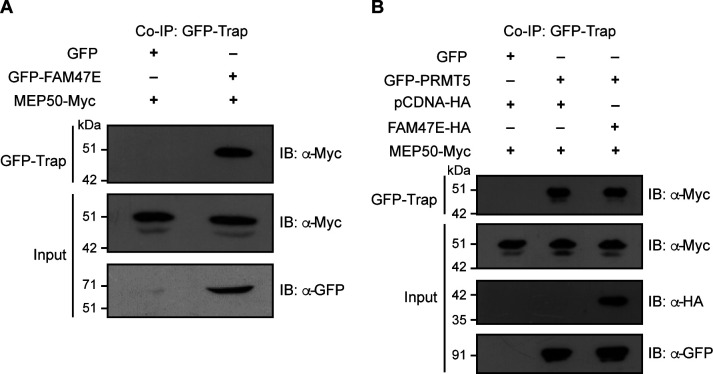
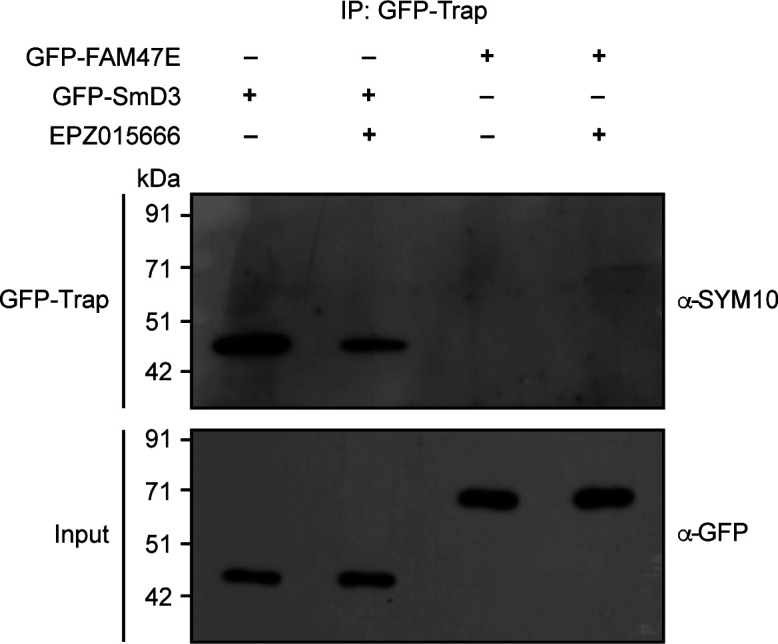
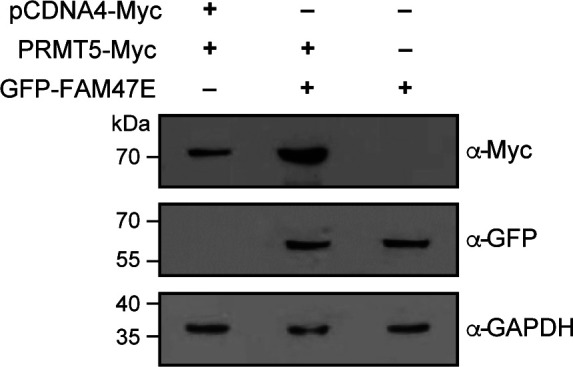
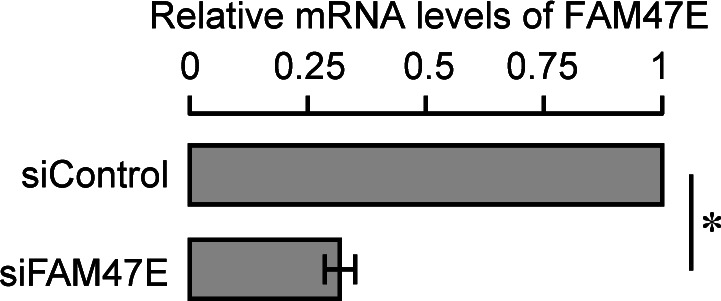
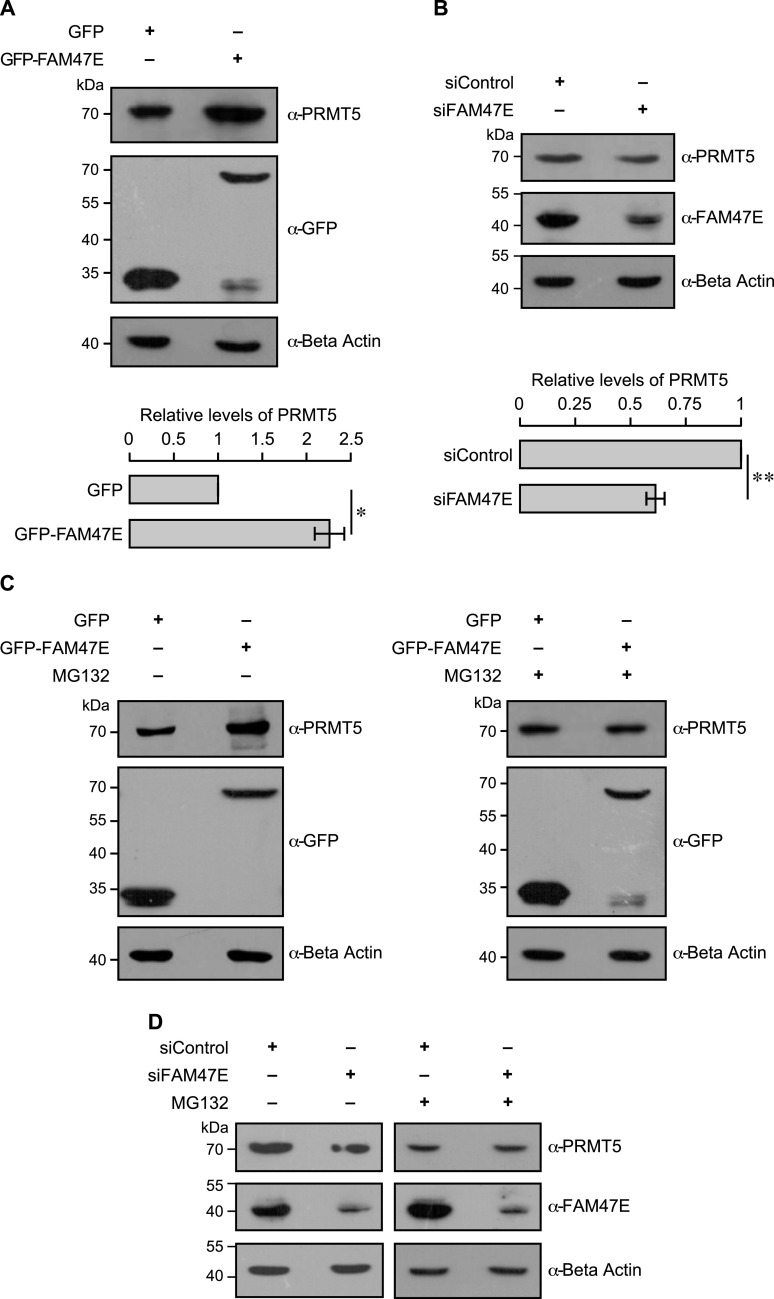
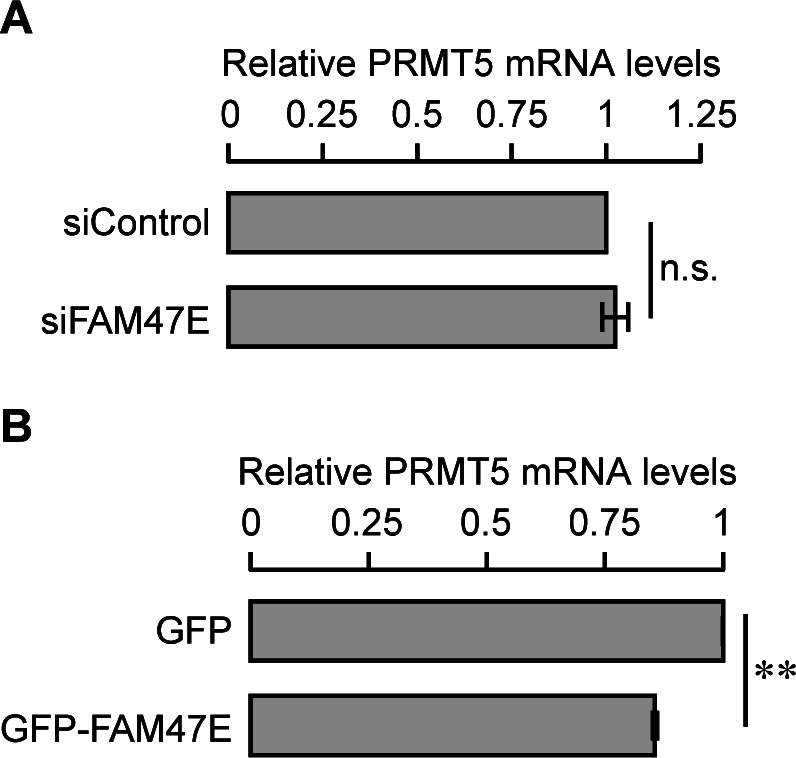
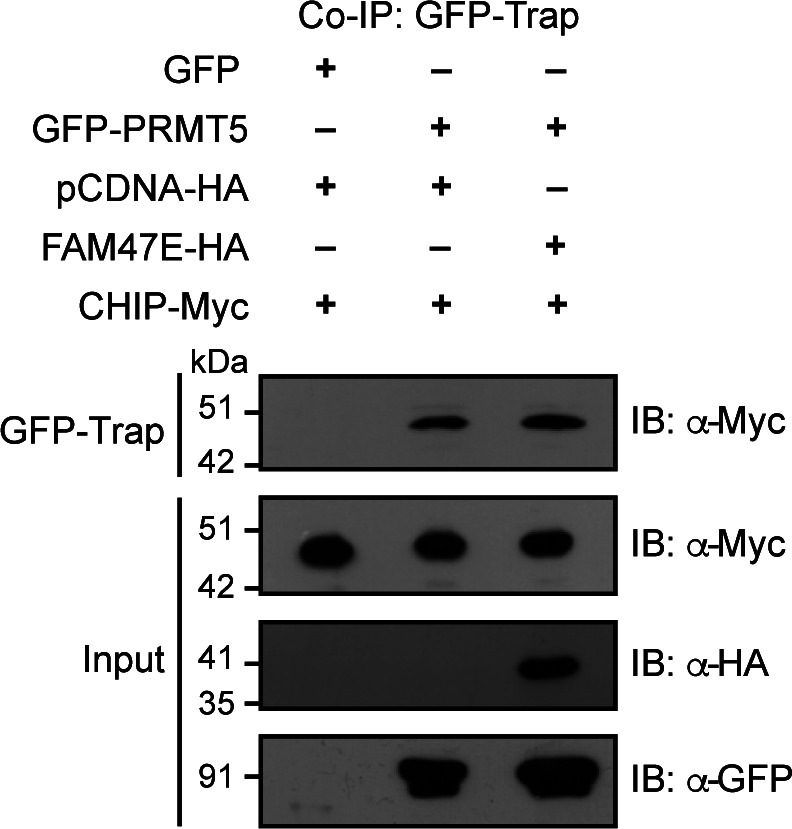
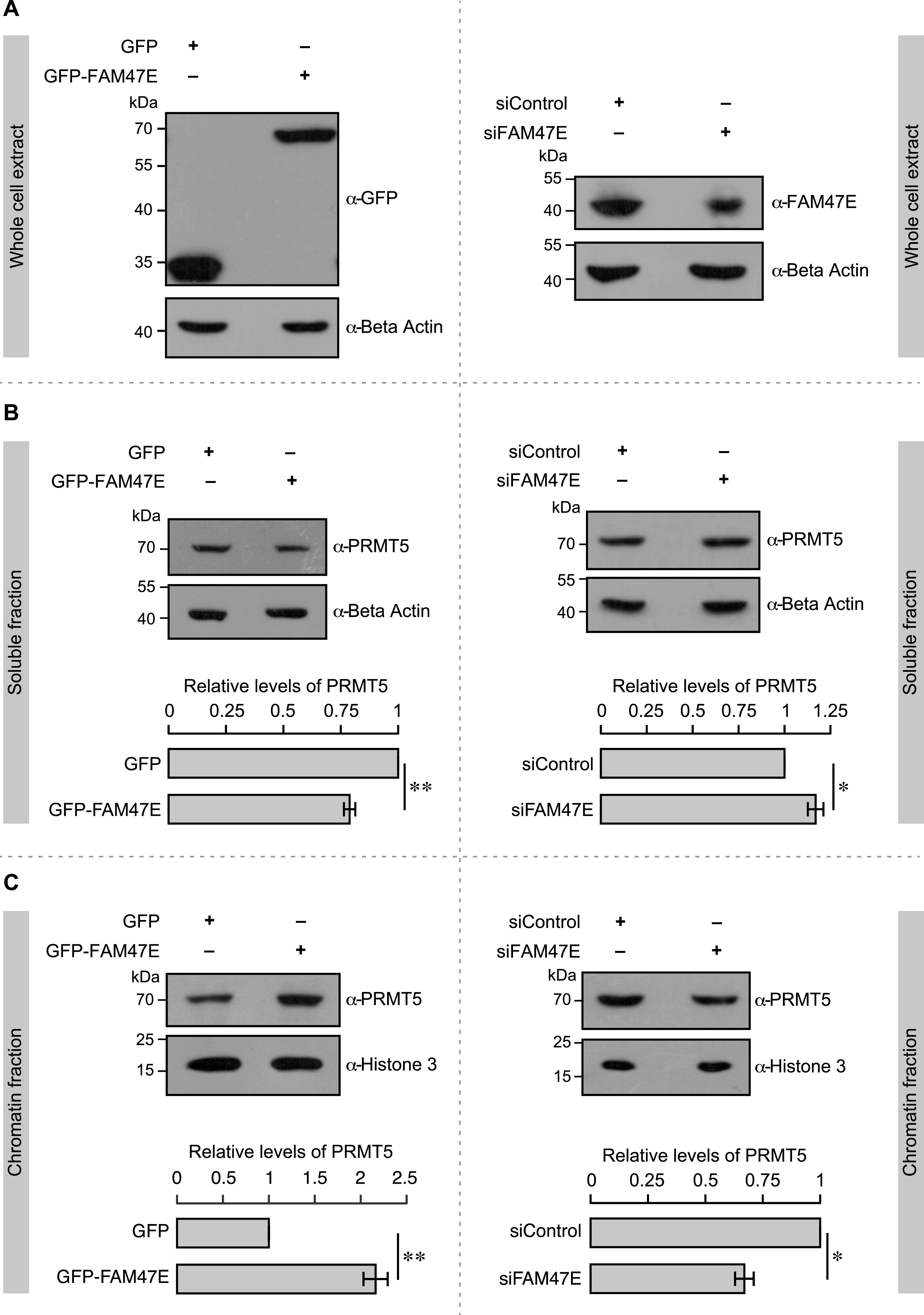
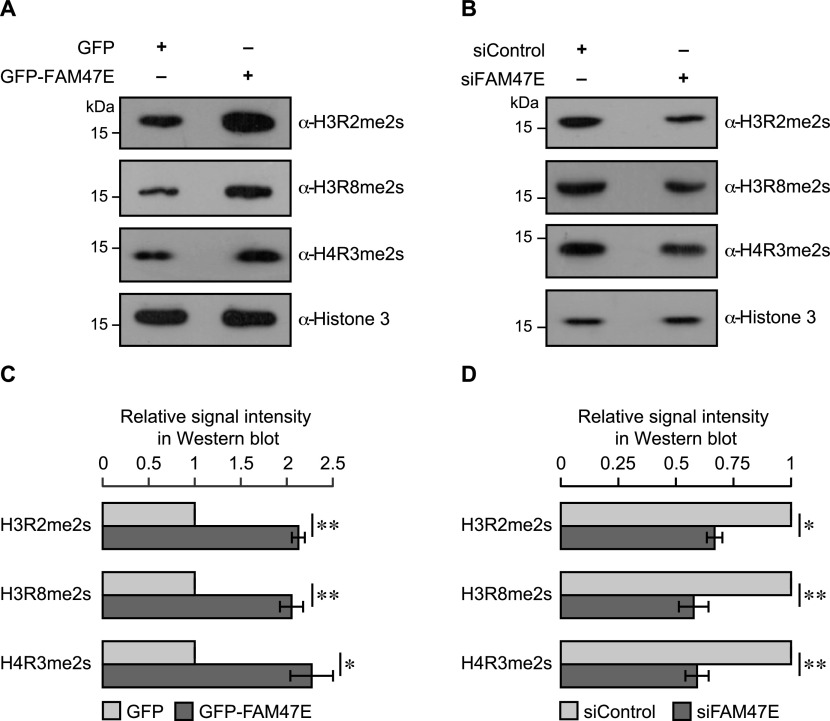
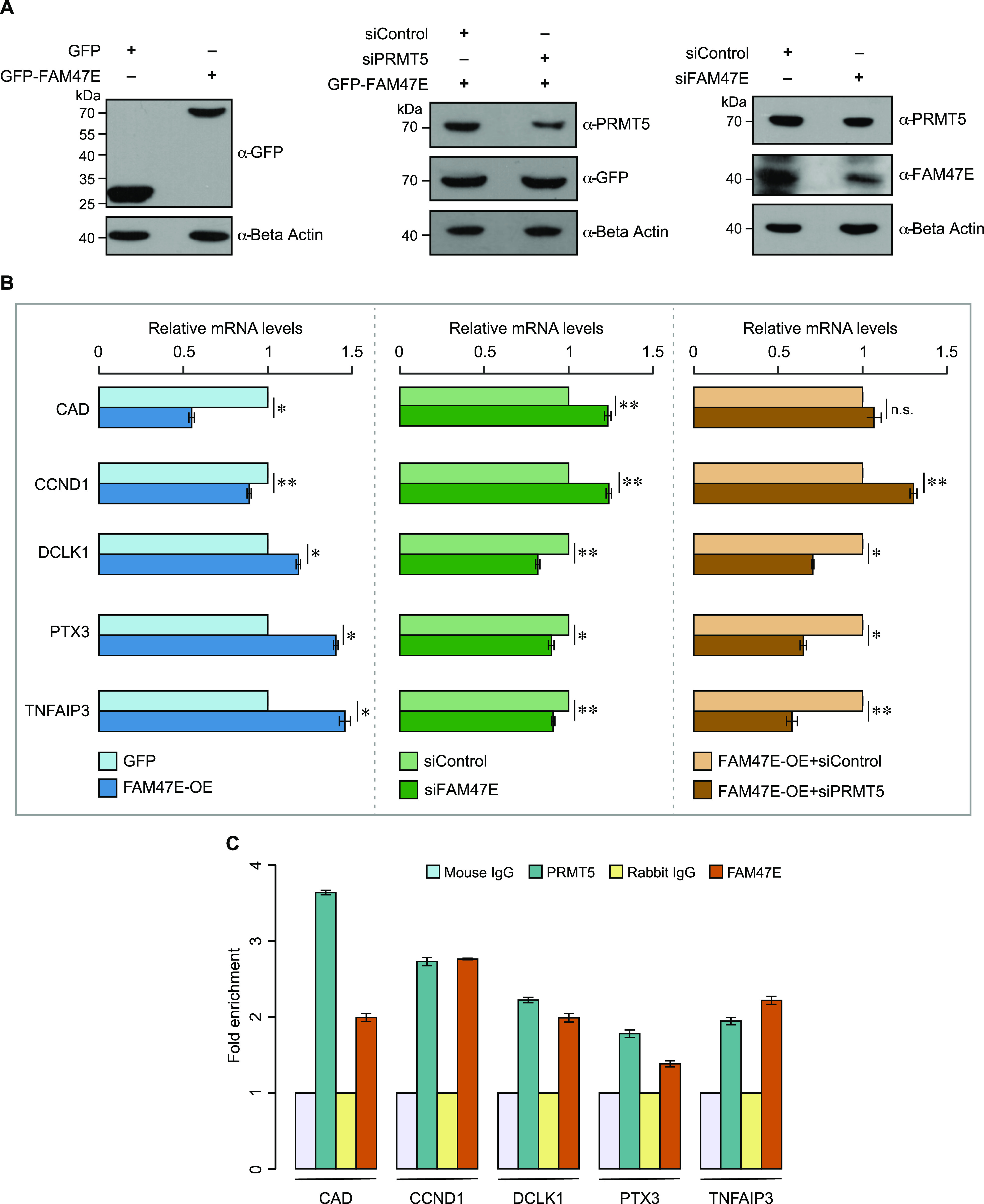
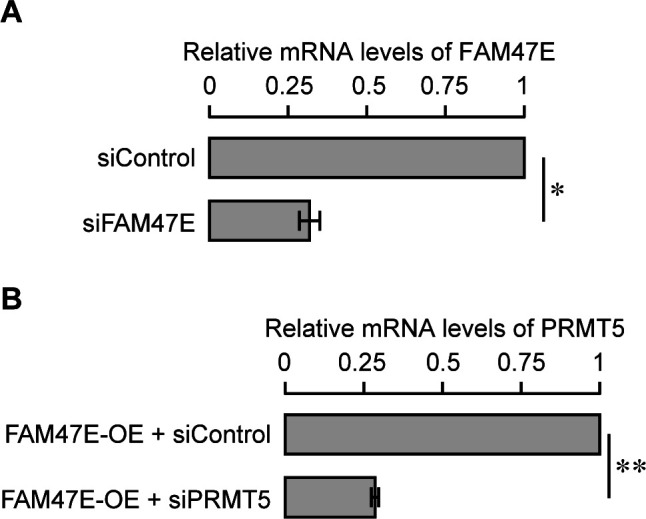
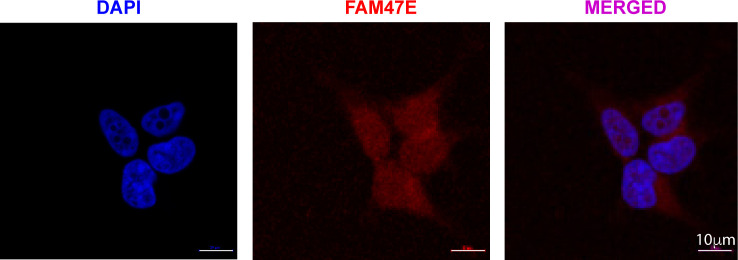
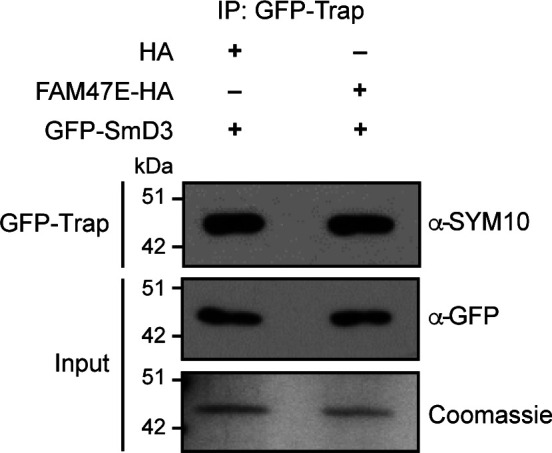
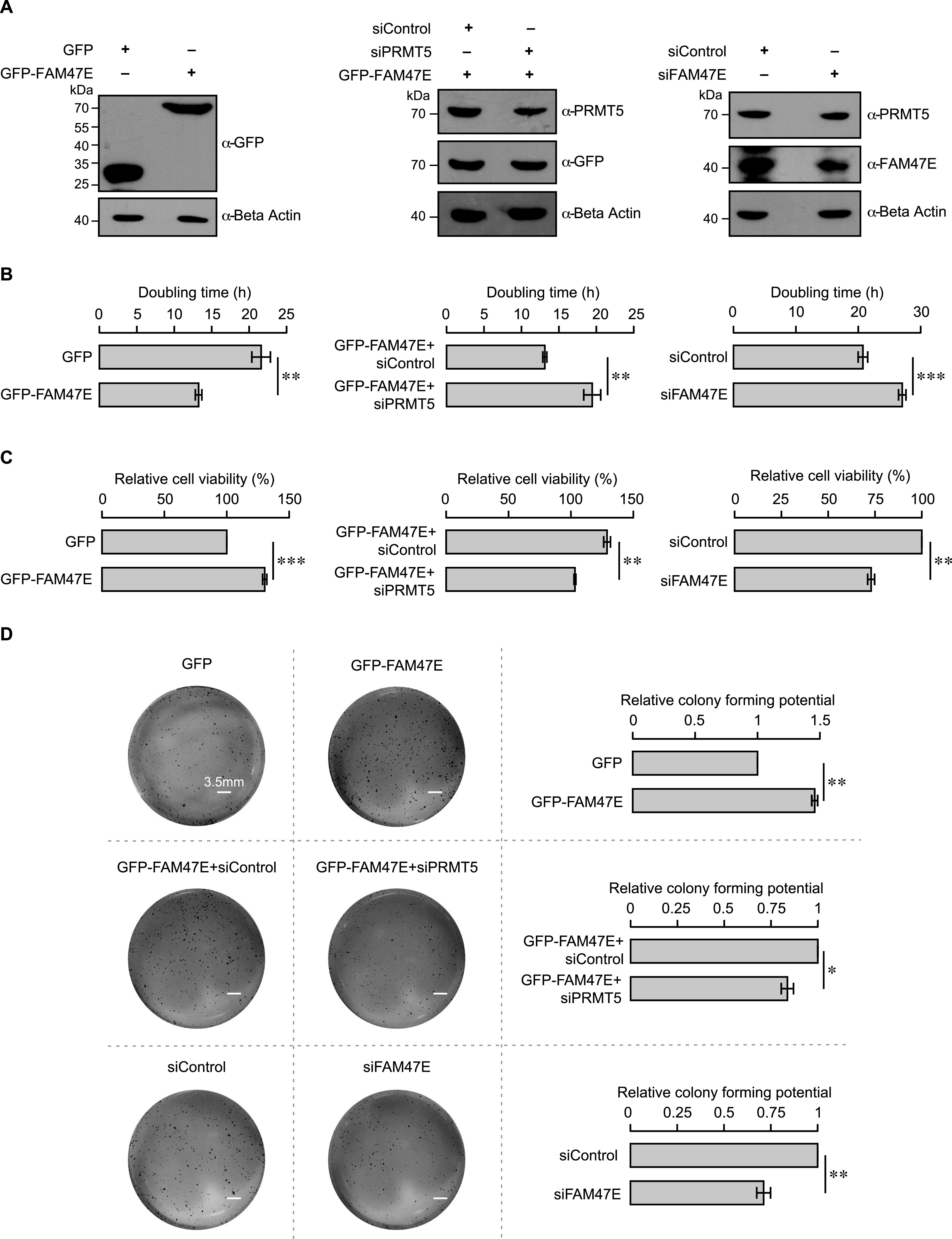
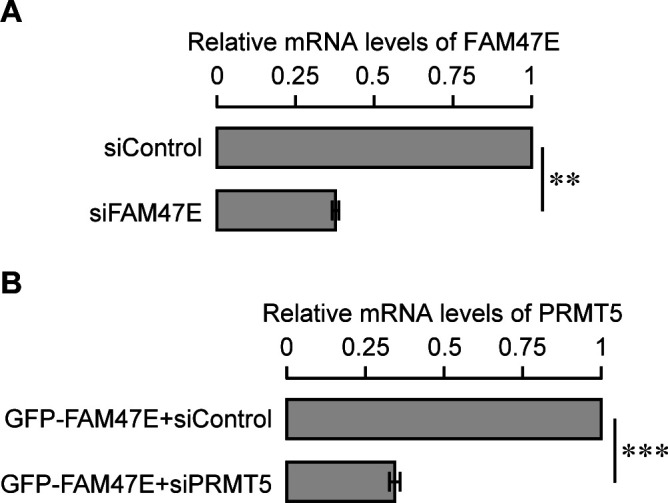
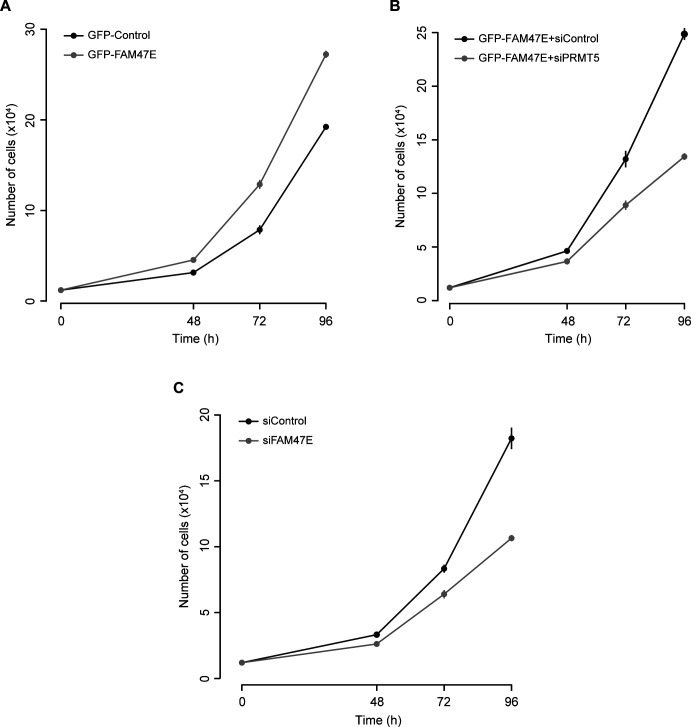
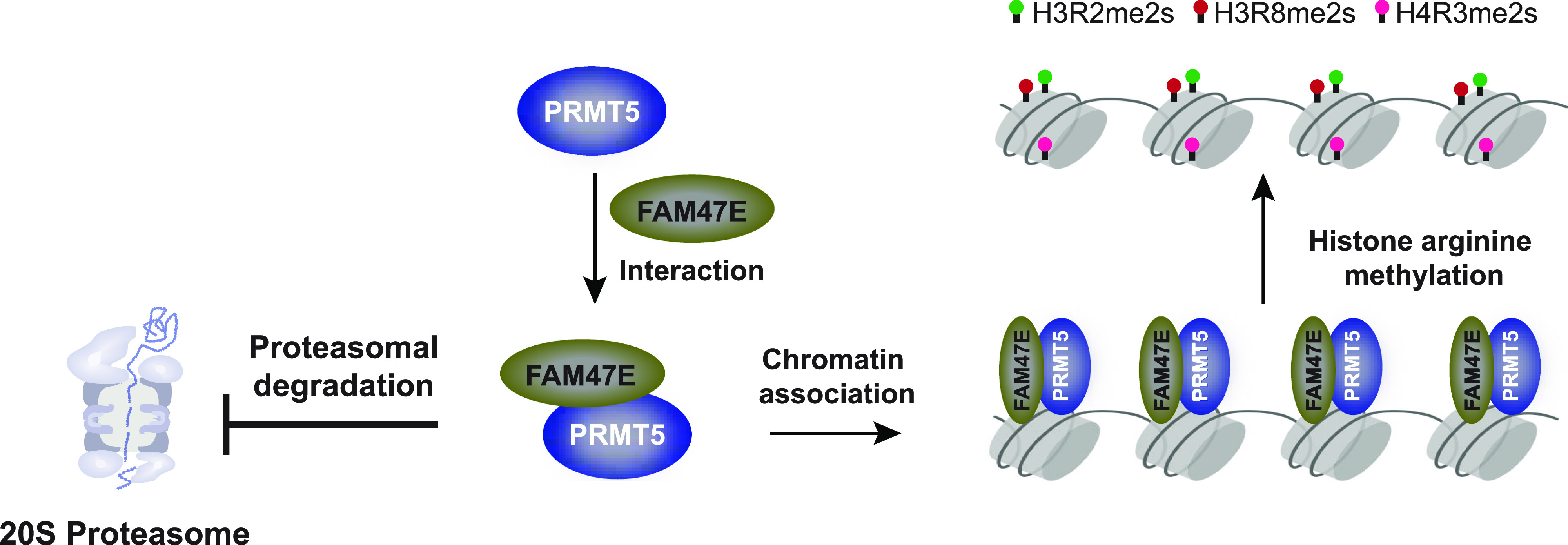
Protein arginine methyltransferase 5 (PRMT5) symmetrically dimethylates arginine residues in various proteins affecting diverse cellular processes such as transcriptional regulation, splicing, DNA repair, differentiation, and cell cycle. Elevated levels of PRMT5 are observed in several types of cancers and are associated with poor clinical outcomes, making PRMT5 an important diagnostic marker and/or therapeutic target for cancers. Here, using yeast two-hybrid screening, followed by immunoprecipitation and pull-down assays, we identify a previously uncharacterized protein, FAM47E, as an interaction partner of PRMT5. We report that FAM47E regulates steady-state levels of PRMT5 by affecting its stability through inhibition of its proteasomal degradation. Importantly, FAM47E enhances the chromatin association and histone methylation activity of PRMT5. The PRMT5-FAM47E interaction affects the regulation of PRMT5 target genes expression and colony-forming capacity of the cells. Taken together, we identify FAM47E as a protein regulator of PRMT5, which promotes the functions of this versatile enzyme. These findings imply that disruption of PRMT5-FAM47E interaction by small molecules might be an alternative strategy to attenuate the oncogenic function(s) of PRMT5.
蛋白质精氨酸甲基转移酶 5(PRMT5)对称地甲基化各种蛋白质中的精氨酸残基,影响转录调控、剪接、DNA 修复、分化和细胞周期等多种细胞过程。几种类型的癌症中观察到 PRMT5 水平升高,并且与不良的临床结果相关,这使得 PRMT5 成为癌症的重要诊断标志物和/或治疗靶标。在这里,我们使用酵母双杂交筛选,然后进行免疫沉淀和下拉测定,鉴定出一个以前未被表征的蛋白质 FAM47E 作为 PRMT5 的相互作用伙伴。我们报告 FAM47E 通过抑制其蛋白酶体降解来影响其稳定性,从而调节 PRMT5 的稳定状态水平。重要的是,FAM47E 增强了 PRMT5 的染色质结合和组蛋白甲基化活性。PRMT5-FAM47E 相互作用影响 PRMT5 靶基因表达的调节和细胞的集落形成能力。总之,我们将 FAM47E 鉴定为 PRMT5 的蛋白质调节剂,它促进了这种多功能酶的功能。这些发现意味着通过小分子破坏 PRMT5-FAM47E 相互作用可能是减弱 PRMT5 致癌功能的另一种策略。