Department of Microbiology, Harvard Medical School, Boston, Massachusetts, USA.
Howard Hughes Medical Institute, Harvard Medical School, Boston, Massachusetts, USA.
mBio. 2021 Jun 29;12(3):e0068221. doi: 10.1128/mBio.00682-21. Epub 2021 Jun 8.
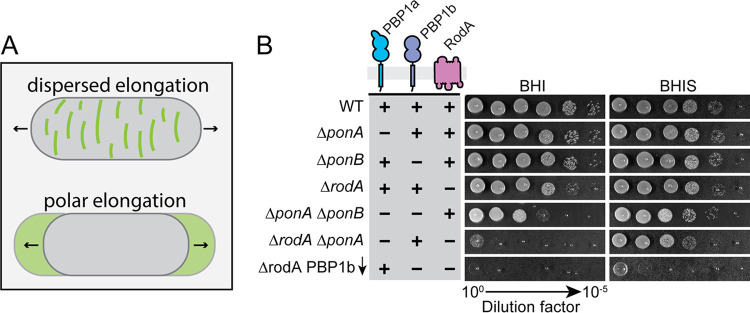
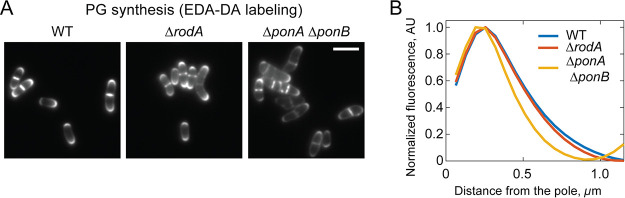
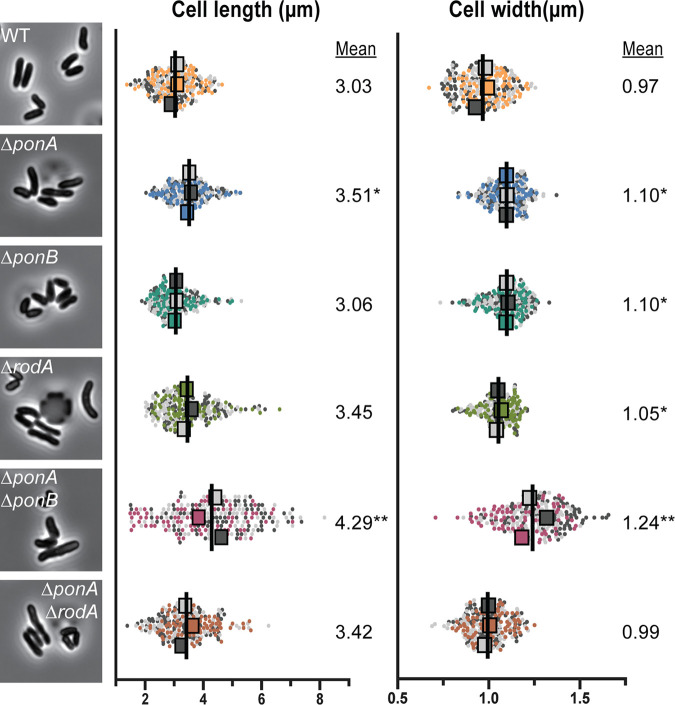
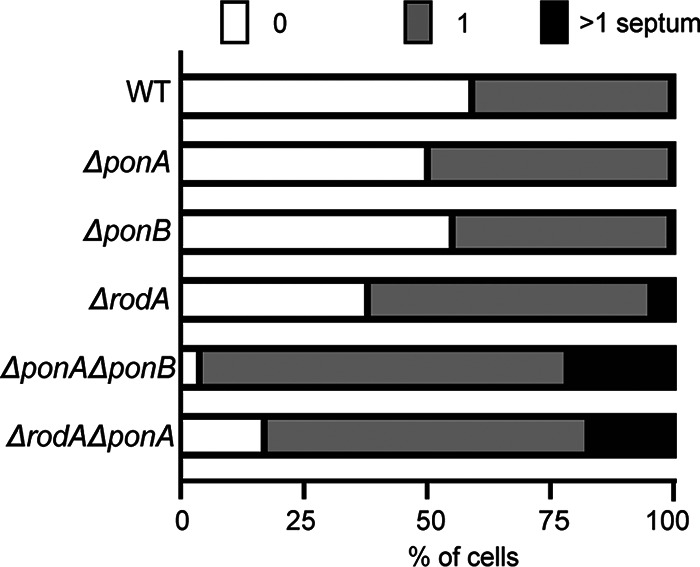
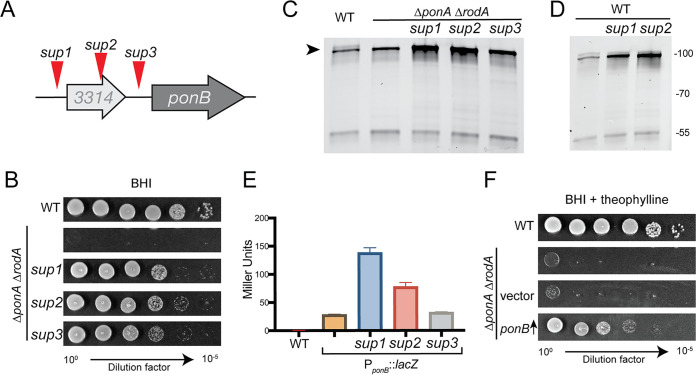
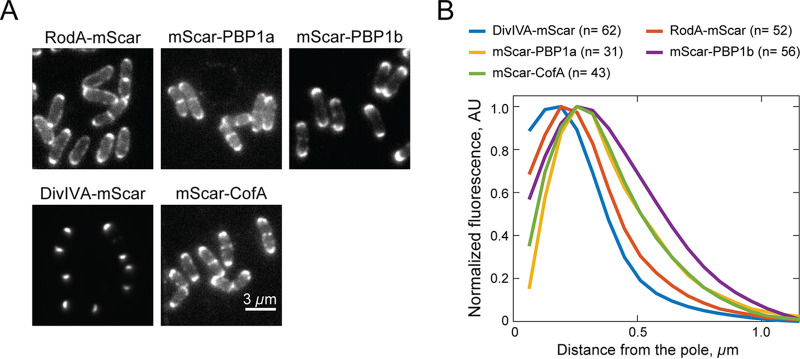
Members of the suborder of bacteria, including major pathogens such as Mycobacterium tuberculosis, grow via the insertion of new cell wall peptidoglycan (PG) material at their poles. This mode of elongation differs from that used by Escherichia coli and other more well-studied model organisms that grow by inserting new PG at dispersed sites along their cell body. Dispersed cell elongation is known to strictly require the SEDS-type PG synthase called RodA, whereas the other major class of PG synthases called class A penicillin-binding proteins (aPBPs) are not required for this mode of growth. Instead, they are thought to be important for maintaining the integrity of the PG matrix in organisms growing by dispersed elongation. In contrast, based on prior genetic studies in M. tuberculosis and related members of the suborder, the aPBPs are widely believed to be essential for polar growth, with RodA being dispensable. However, polar growth has not been directly assessed in mycobacterial or corynebacterial mutants lacking aPBP-type PG synthases. We therefore investigated the relative roles of aPBPs and RodA in polar growth using Corynebacterium glutamicum as a model member of . Notably, we discovered that the aPBPs are dispensable for polar growth and that this growth mode can be mediated by either an aPBP-type or a SEDS-type enzyme functioning as the sole elongation PG synthase. Thus, our results reveal that the mechanism of polar elongation is fundamentally flexible and, unlike dispersed elongation, can be effectively mediated in C. glutamicum by either a SEDS-bPBP or an aPBP-type synthase. The suborder includes a number of major bacterial pathogens. These organisms grow by polar extension unlike most well-studied model bacteria, which grow by inserting wall material at dispersed sites along their length. A better understanding of polar growth promises to uncover new avenues for targeting mycobacterial and corynebacterial infections. Here, we investigated the roles of the different classes of cell wall synthases for polar growth using Corynebacterium glutamicum as a model. We discovered that the polar growth mechanism is surprisingly flexible in this organism and, unlike dispersed synthesis, can function using either of the two known types of cell wall synthase enzymes.
该亚目的细菌成员,包括结核分枝杆菌等主要病原体,通过在其极部长出新的细胞壁肽聚糖(PG)物质来生长。这种伸长模式与大肠杆菌和其他研究更为充分的模式生物不同,后者通过在其细胞体上分散的位置插入新的 PG 来生长。已知分散的细胞伸长严格需要称为 RodA 的 SEDS 型 PG 合酶,而其他主要的 PG 合酶类称为 A 类青霉素结合蛋白(aPBPs),则不需要这种生长模式。相反,它们被认为对于通过分散伸长生长的生物体中 PG 基质的完整性很重要。相比之下,基于先前在结核分枝杆菌和该亚目相关成员中的遗传研究,普遍认为 aPBPs 对于极性生长是必需的,而 RodA 则是可有可无的。然而,在缺乏 aPBP 型 PG 合酶的分枝杆菌或棒状杆菌突变体中,尚未直接评估极性生长。因此,我们使用谷氨酸棒状杆菌作为 的模型成员,研究了 aPBPs 和 RodA 在极性生长中的相对作用。值得注意的是,我们发现 aPBPs 对于极性生长是可有可无的,并且这种生长模式可以由作为唯一伸长 PG 合酶的 aPBP 型或 SEDS 型酶介导。因此,我们的结果表明,极性伸长的机制从根本上是灵活的,与分散伸长不同,在谷氨酸棒状杆菌中可以有效地由 SEDS-bPBP 或 aPBP 型合酶介导。该亚目包括许多主要的细菌病原体。这些生物体通过极性延伸生长,与大多数研究充分的模式细菌不同,后者通过在其长度上分散的位置插入壁材料来生长。更好地了解极性生长有望为靶向分枝杆菌和棒状杆菌感染开辟新途径。在这里,我们使用谷氨酸棒状杆菌作为模型,研究了不同细胞壁合酶类对极性生长的作用。我们发现,在这种生物体中,极性生长机制出人意料地灵活,与分散合成不同,它可以使用两种已知类型的细胞壁合酶之一发挥作用。