Structural Biology Division, The Walter and Eliza Hall Institute of Medical Research, Parkville, Victoria, Australia; Department of Medical Biology, University of Melbourne, Parkville, Victoria, Australia.
Structural Biology Division, The Walter and Eliza Hall Institute of Medical Research, Parkville, Victoria, Australia.
J Biol Chem. 2021 Jul;297(1):100900. doi: 10.1016/j.jbc.2021.100900. Epub 2021 Jun 19.
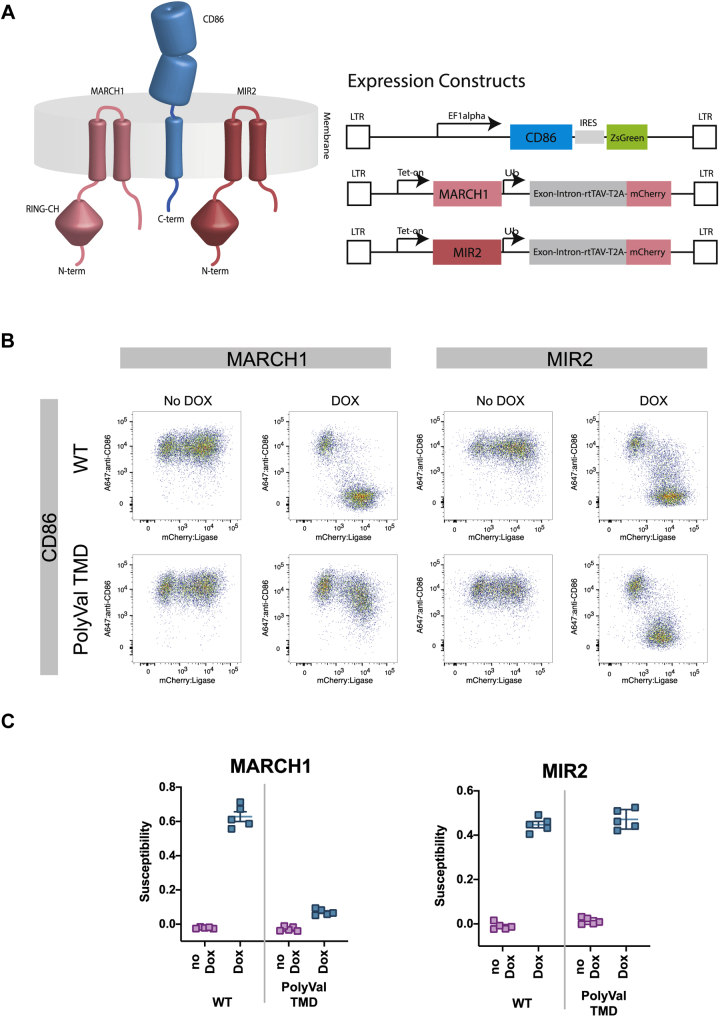
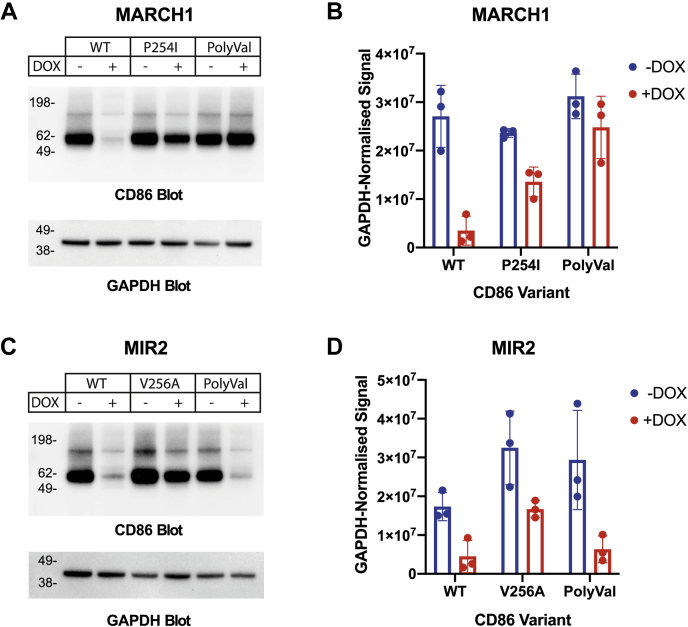
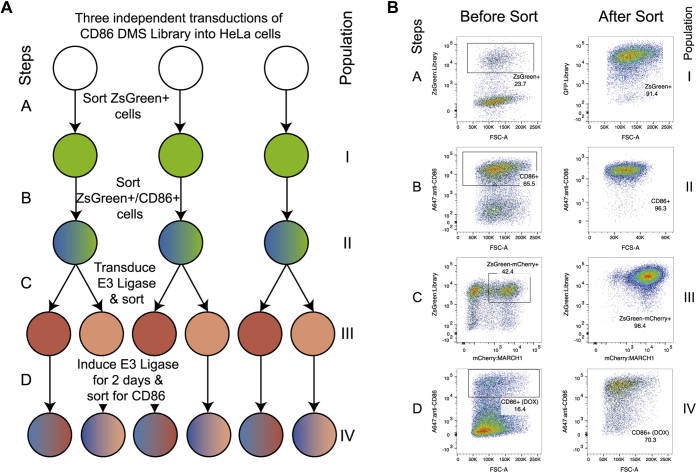
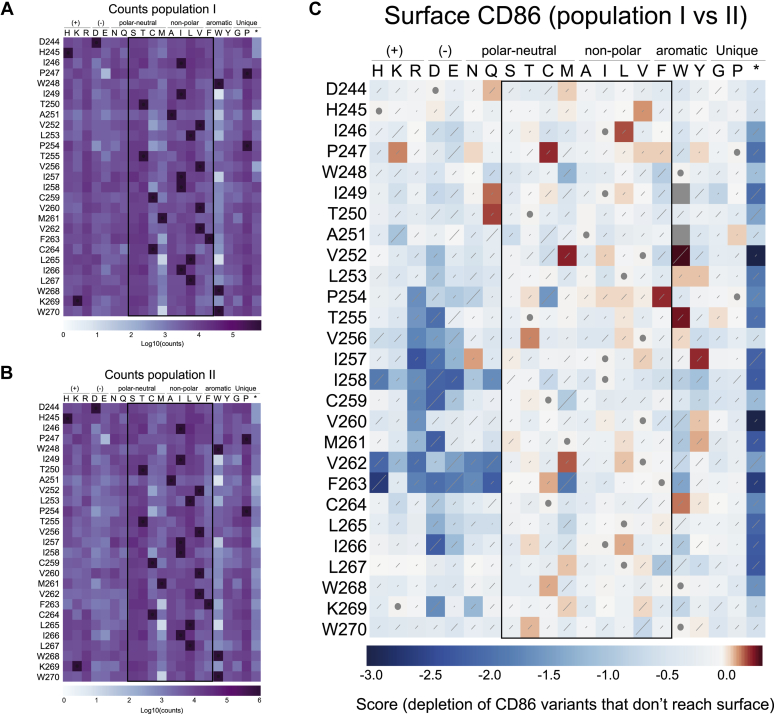
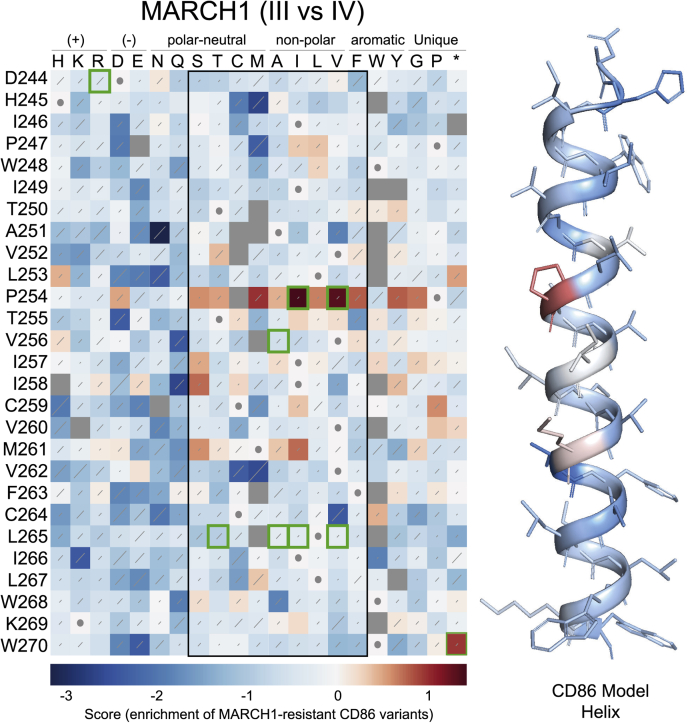
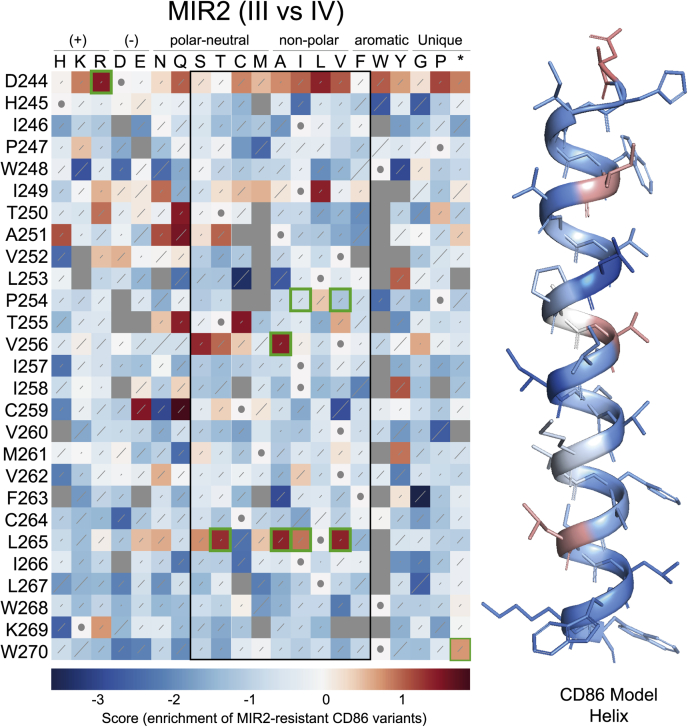
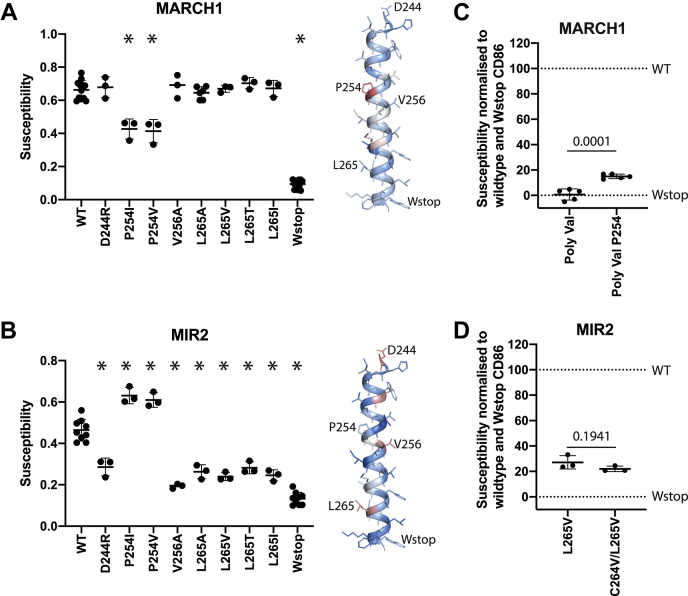
Immune-stimulatory ligands, such as major histocompatibility complex molecules and the T-cell costimulatory ligand CD86, are central to productive immunity. Endogenous mammalian membrane-associated RING-CHs (MARCH) act on these and other targets to regulate antigen presentation and activation of adaptive immunity, whereas virus-encoded homologs target the same molecules to evade immune responses. Substrate specificity is encoded in or near the membrane-embedded domains of MARCHs and the proteins they regulate, but the exact sequences that distinguish substrates from nonsubstrates are poorly understood. Here, we examined the requirements for recognition of the costimulatory ligand CD86 by two different MARCH-family proteins, human MARCH1 and Kaposi's sarcoma herpesvirus modulator of immune recognition 2 (MIR2), using deep mutational scanning. We identified a highly specific recognition surface in the hydrophobic core of the CD86 transmembrane (TM) domain (TMD) that is required for recognition by MARCH1 and prominently features a proline at position 254. In contrast, MIR2 requires no specific sequences in the CD86 TMD but relies primarily on an aspartic acid at position 244 in the CD86 extracellular juxtamembrane region. Surprisingly, MIR2 recognized CD86 with a TMD composed entirely of valine, whereas many different single amino acid substitutions in the context of the native TM sequence conferred MIR2 resistance. These results show that the human and viral proteins evolved completely different recognition modes for the same substrate. That some TM sequences are incompatible with MIR2 activity, even when no specific recognition motif is required, suggests a more complicated mechanism of immune modulation via CD86 than was previously appreciated.
免疫刺激配体,如主要组织相容性复合体分子和 T 细胞共刺激配体 CD86,是产生免疫的核心。内源性哺乳动物膜相关 RING-CH(MARCH)作用于这些和其他靶标,以调节抗原呈递和适应性免疫的激活,而病毒编码的同源物则针对相同的分子来逃避免疫反应。MARCH 和它们调节的蛋白质的底物特异性编码在膜嵌入结构域内或附近,但区分底物和非底物的确切序列知之甚少。在这里,我们使用深度突变扫描研究了两种不同的 MARCH 家族蛋白,人 MARCH1 和卡波济肉瘤疱疹病毒免疫识别调节剂 2(MIR2)识别共刺激配体 CD86 的要求。我们在 CD86 跨膜(TM)域(TMD)的疏水区鉴定了一个高度特异性的识别表面,该表面对于 MARCH1 和 Kaposi 肉瘤疱疹病毒免疫识别调节剂 2(MIR2)的识别是必需的,其特征是在位置 254 处有一个脯氨酸。相比之下,MIR2 在 CD86 TMD 中不需要特定的序列,但主要依赖于 CD86 细胞外近膜区的位置 244 处的天冬氨酸。令人惊讶的是,MIR2 识别 CD86 的 TMD 完全由缬氨酸组成,而在天然 TM 序列的背景下,许多不同的单个氨基酸取代赋予了 MIR2 抗性。这些结果表明,人类和病毒蛋白为同一底物进化出了完全不同的识别模式。一些 TM 序列与 MIR2 活性不兼容,即使不需要特定的识别模体,这表明通过 CD86 进行免疫调节的机制比以前认为的更为复杂。