Schut Gerrit J, Haja Dominik K, Feng Xiang, Poole Farris L, Li Huilin, Adams Michael W W
Department of Biochemistry and Molecular Biology, University of Georgia, Athens, GA, United States.
Department of Structural Biology, Van Andel Institute, Grand Rapids, MI, United States.
Front Microbiol. 2022 Jul 8;13:946711. doi: 10.3389/fmicb.2022.946711. eCollection 2022.
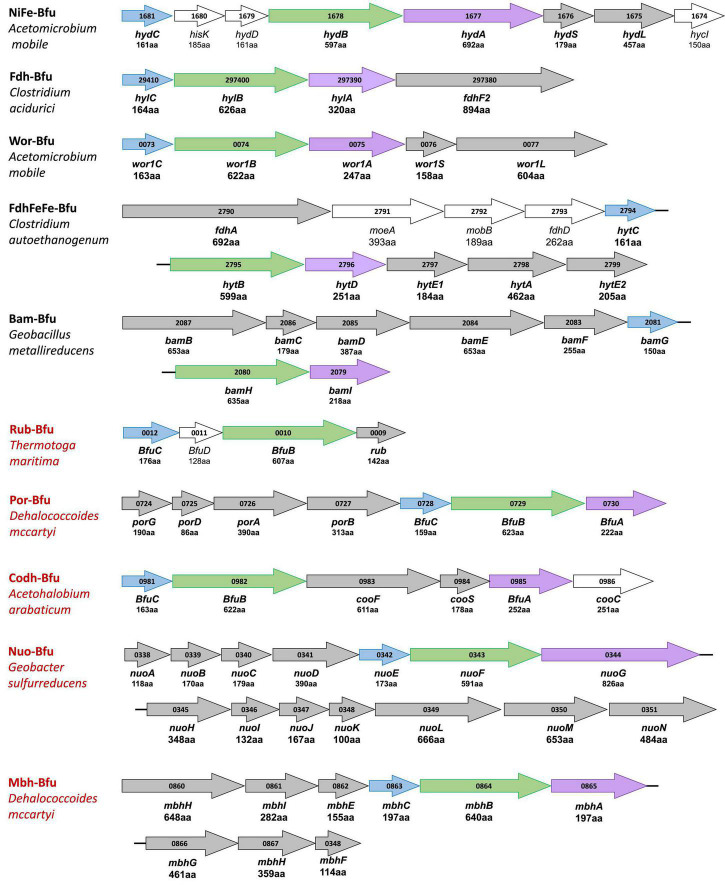
Microorganisms utilize electron bifurcating enzymes in metabolic pathways to carry out thermodynamically unfavorable reactions. Bifurcating FeFe-hydrogenases (HydABC) reversibly oxidize NADH (E'∼-280 mV, under physiological conditions) and reduce protons to H gas (E°'-414 mV) by coupling this endergonic reaction to the exergonic reduction of protons by reduced ferredoxin (Fd) (E'∼-500 mV). We show here that HydABC homologs are surprisingly ubiquitous in the microbial world and are represented by 57 phylogenetically distinct clades but only about half are FeFe-hydrogenases. The others have replaced the hydrogenase domain with another oxidoreductase domain or they contain additional subunits, both of which enable various third reactions to be reversibly coupled to NAD and Fd reduction. We hypothesize that all of these enzymes carry out electron bifurcation and that their third substrates can include hydrogen peroxide, pyruvate, carbon monoxide, aldehydes, aryl-CoA thioesters, NADP, cofactor F, formate, and quinones, as well as many yet to be discovered. Some of the enzymes are proposed to be integral membrane-bound proton-translocating complexes. These different functionalities are associated with phylogenetically distinct clades and in many cases with specific microbial phyla. We propose that this new and abundant class of electron bifurcating enzyme be referred to as the Bfu family whose defining feature is a conserved bifurcating BfuBC core. This core contains FMN and six iron sulfur clusters and it interacts directly with ferredoxin (Fd) and NAD(H). Electrons to or from the third substrate are fed into the BfuBC core BfuA. The other three known families of electron bifurcating enzyme (abbreviated as Nfn, EtfAB, and HdrA) contain a special FAD that bifurcates electrons to high and low potential pathways. The Bfu family are proposed to use a different electron bifurcation mechanism that involves a combination of FMN and three adjacent iron sulfur clusters, including a novel [2Fe-2S] cluster with pentacoordinate and partial non-Cys coordination. The absolute conservation of the redox cofactors of BfuBC in all members of the Bfu enzyme family indicate they have the same non-canonical mechanism to bifurcate electrons. A hypothetical catalytic mechanism is proposed as a basis for future spectroscopic analyses of Bfu family members.
微生物在代谢途径中利用电子分叉酶来进行热力学上不利的反应。分叉型铁铁氢化酶(HydABC)可逆地氧化NADH(在生理条件下E' ∼ -280 mV),并通过将这个吸能反应与还原型铁氧化还原蛋白(Fd)(E' ∼ -500 mV)对质子的放能还原相偶联,将质子还原为氢气(E°' -414 mV)。我们在此表明,HydABC同源物在微生物界惊人地普遍存在,由57个系统发育上不同的进化枝代表,但只有约一半是铁铁氢化酶。其他的则用另一个氧化还原酶结构域取代了氢化酶结构域,或者它们含有额外的亚基,这两者都能使各种第三个反应与NAD和Fd的还原可逆地偶联。我们推测所有这些酶都进行电子分叉,并且它们的第三个底物可以包括过氧化氢、丙酮酸、一氧化碳、醛、芳基-CoA硫酯、NADP、辅因子F、甲酸和醌,以及许多尚未被发现的物质。一些酶被认为是整合膜结合的质子转运复合物。这些不同的功能与系统发育上不同的进化枝相关,并且在许多情况下与特定的微生物门类相关。我们建议将这类新的且丰富的电子分叉酶称为Bfu家族,其定义特征是保守的分叉BfuBC核心。这个核心包含FMN和六个铁硫簇,并且它直接与铁氧化还原蛋白(Fd)和NAD(H)相互作用。进出第三个底物的电子被输入到BfuBC核心与BfuA中。其他三个已知的电子分叉酶家族(简称为Nfn、EtfAB和HdrA)包含一种特殊的FAD,它将电子分叉到高电位和低电位途径。Bfu家族被认为使用一种不同地电子分叉机制,该机制涉及FMN和三个相邻的铁硫簇的组合,包括一个具有五配位和部分非半胱氨酸配位的新型[2Fe-2S]簇。BfuBC的氧化还原辅因子在Bfu酶家族的所有成员中绝对保守,这表明它们具有相同的非经典电子分叉机制。提出了一种假设的催化机制作为未来对Bfu家族成员进行光谱分析的基础。