Gisriel Christopher J, Cardona Tanai, Bryant Donald A, Brudvig Gary W
Department of Chemistry, Yale University, New Haven, CT 06520, USA.
Department of Life Sciences, Imperial College London, London SW7 2AZ, UK.
Microorganisms. 2022 Jun 22;10(7):1270. doi: 10.3390/microorganisms10071270.
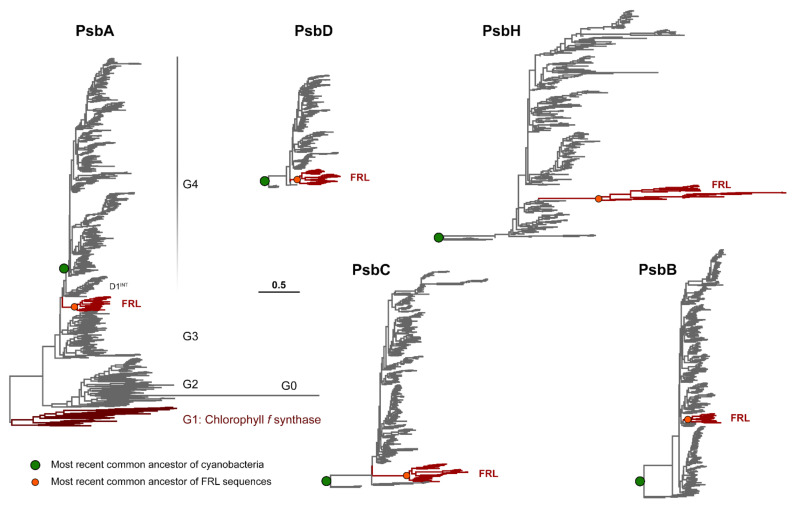
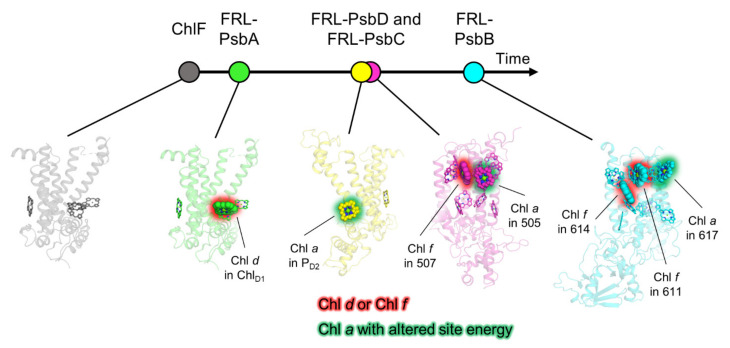
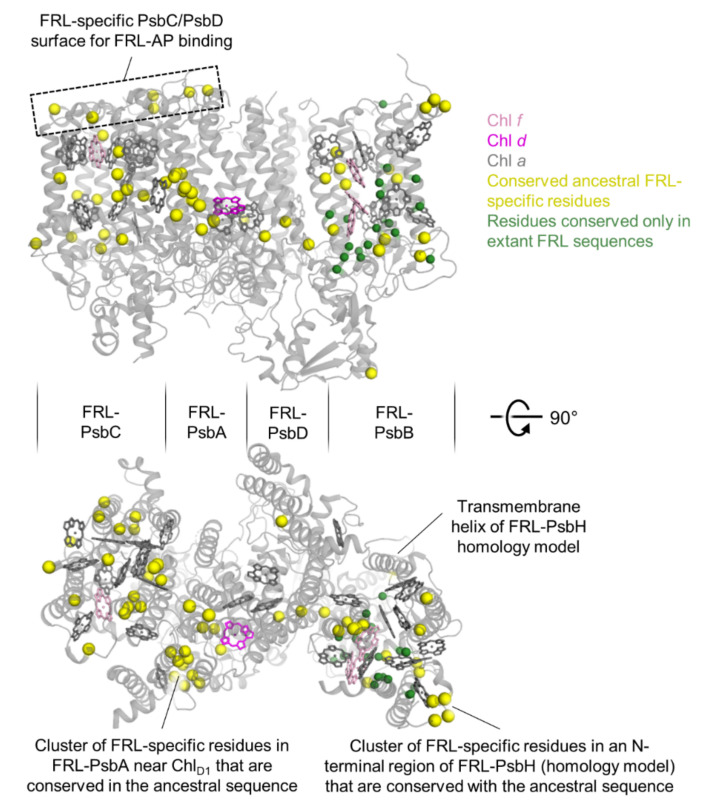
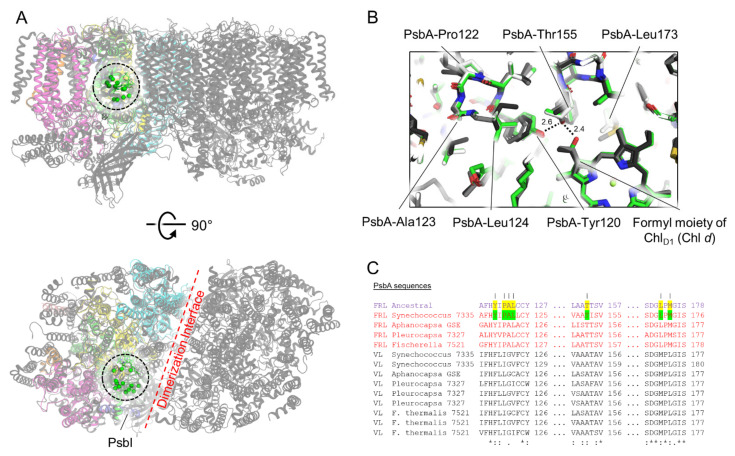
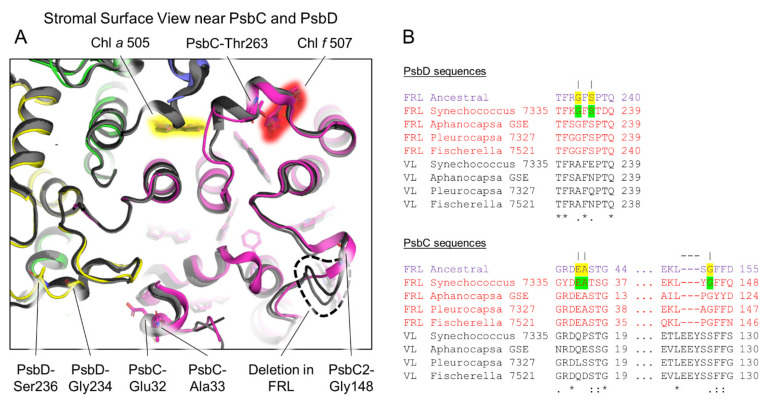
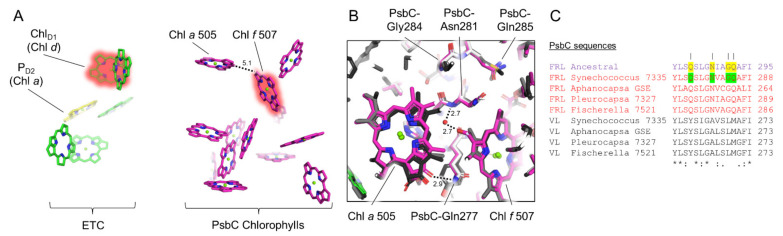
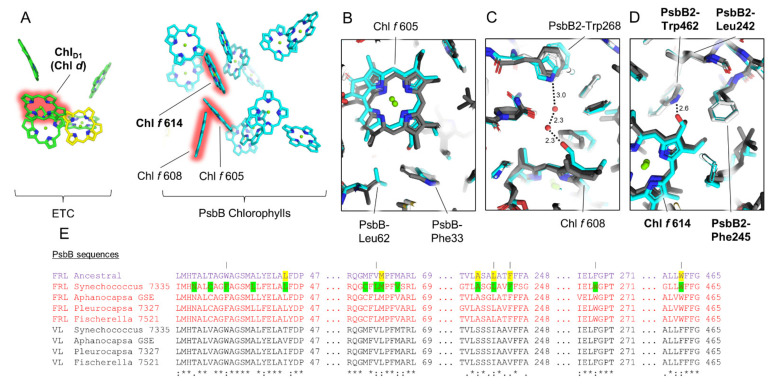
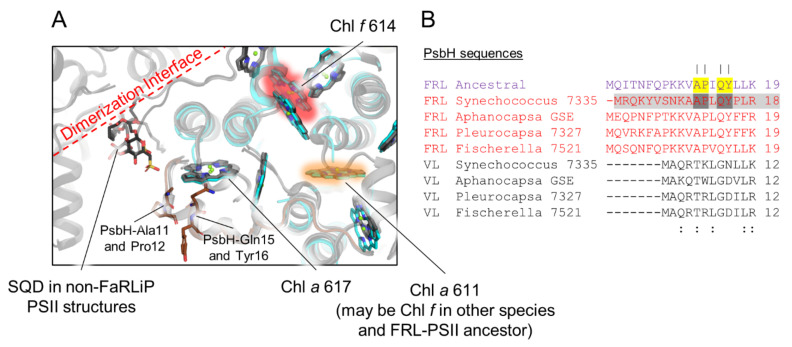
Cyanobacteria are major contributors to global carbon fixation and primarily use visible light (400-700 nm) to drive oxygenic photosynthesis. When shifted into environments where visible light is attenuated, a small, but highly diverse and widespread number of cyanobacteria can express modified pigments and paralogous versions of photosystem subunits and phycobiliproteins that confer far-red light (FRL) absorbance (700-800 nm), a process termed far-red light photoacclimation, or FaRLiP. During FaRLiP, alternate photosystem II (PSII) subunits enable the complex to bind chlorophylls and , which absorb at lower energy than chlorophyll but still support water oxidation. How the FaRLiP response arose remains poorly studied. Here, we report ancestral sequence reconstruction and structure-based molecular evolutionary studies of the FRL-specific subunits of FRL-PSII. We show that the duplications leading to the origin of two PsbA (D1) paralogs required to make chlorophyll and to bind chlorophyll in water-splitting FRL-PSII are likely the first to have occurred prior to the diversification of extant cyanobacteria. These duplications were followed by those leading to alternative PsbC (CP43) and PsbD (D2) subunits, occurring early during the diversification of cyanobacteria, and culminating with those leading to PsbB (CP47) and PsbH paralogs coincident with the radiation of the major groups. We show that the origin of FRL-PSII required the accumulation of a relatively small number of amino acid changes and that the ancestral FRL-PSII likely contained a chlorophyll molecule in the electron transfer chain, two chlorophyll molecules in the antenna subunits at equivalent positions, and three chlorophyll molecules whose site energies were altered. The results suggest a minimal model for engineering far-red light absorbance into plant PSII for biotechnological applications.
蓝细菌是全球碳固定的主要贡献者,主要利用可见光(400 - 700纳米)驱动产氧光合作用。当转移到可见光衰减的环境中时,一小部分但种类繁多且广泛分布的蓝细菌能够表达修饰的色素以及光系统亚基和藻胆蛋白的旁系同源版本,这些赋予了远红光(FRL)吸收能力(700 - 800纳米),这一过程称为远红光光适应,或FaRLiP。在FaRLiP过程中,交替的光系统II(PSII)亚基使该复合体能够结合叶绿素 和 ,它们比叶绿素 吸收能量更低,但仍能支持水氧化。FaRLiP反应是如何产生的,目前仍研究不足。在这里,我们报告了FRL - PSII的FRL特异性亚基的祖先序列重建和基于结构的分子进化研究。我们表明,导致在水裂解FRL - PSII中结合叶绿素 和 所需的两个PsbA(D1)旁系同源物起源的复制事件,很可能是在现存蓝细菌多样化之前首次发生的。这些复制之后是导致替代的PsbC(CP43)和PsbD(D2)亚基的复制,发生在蓝细菌多样化的早期,并随着导致PsbB(CP47)和PsbH旁系同源物的复制达到顶峰,这与主要类群的辐射同时发生。我们表明,FRL - PSII的起源需要积累相对少量的氨基酸变化,并且祖先的FRL - PSII可能在电子传递链中含有一个叶绿素 分子,在天线亚基的等效位置含有两个叶绿素 分子,以及三个位点能量发生改变的叶绿素 分子。这些结果为将远红光吸收工程引入植物PSII以用于生物技术应用提出了一个最小模型。