Department of Plant and Microbial Biology, University of California Berkeley, Berkeley, CA 94720, U.S.A.
Innovative Genomics Institute, University of California Berkeley, 2151 Berkeley Way, Berkeley, CA 94720, U.S.A.
Mol Plant Microbe Interact. 2023 Jul;36(7):434-446. doi: 10.1094/MPMI-07-22-0154-R. Epub 2023 Aug 14.
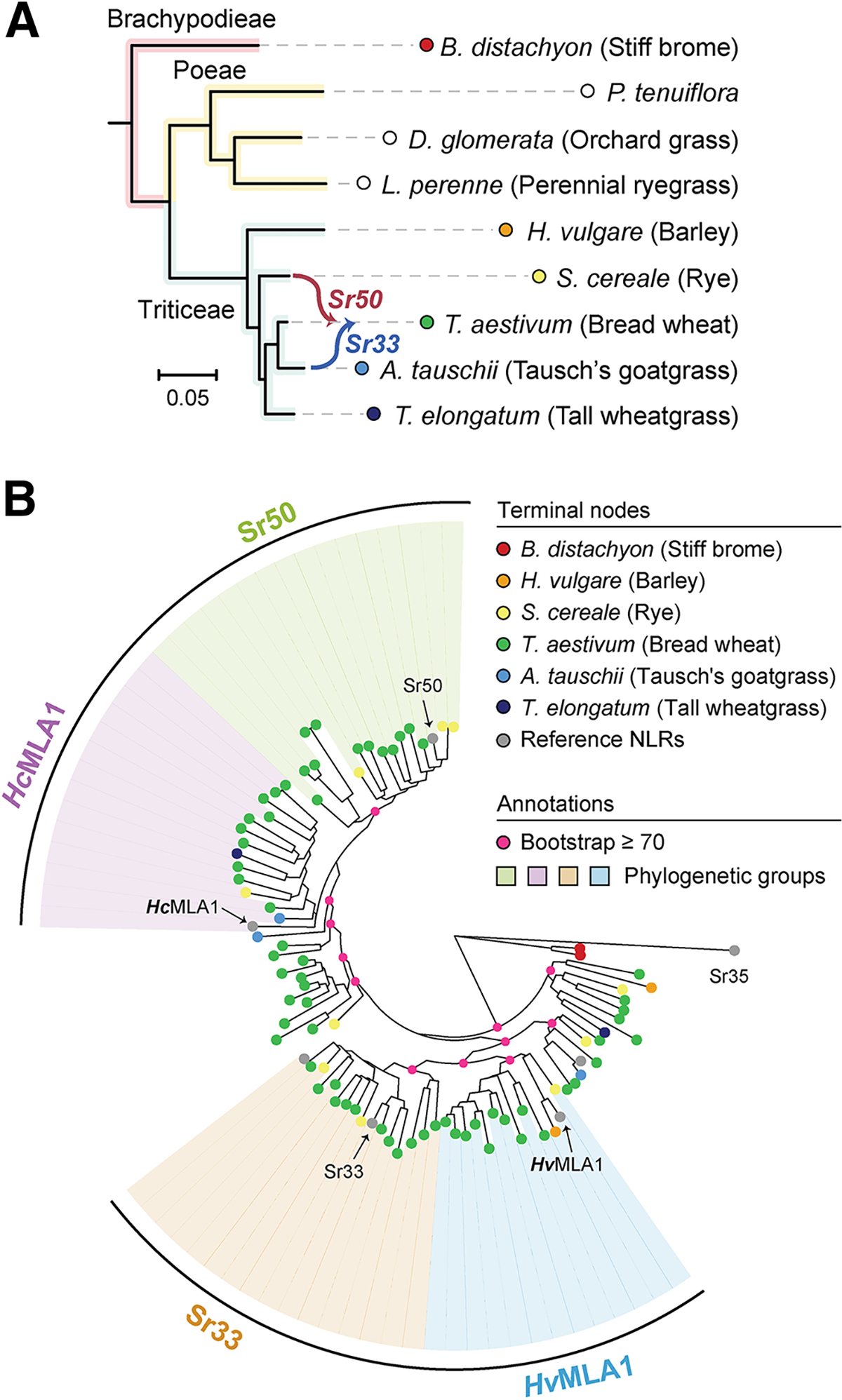
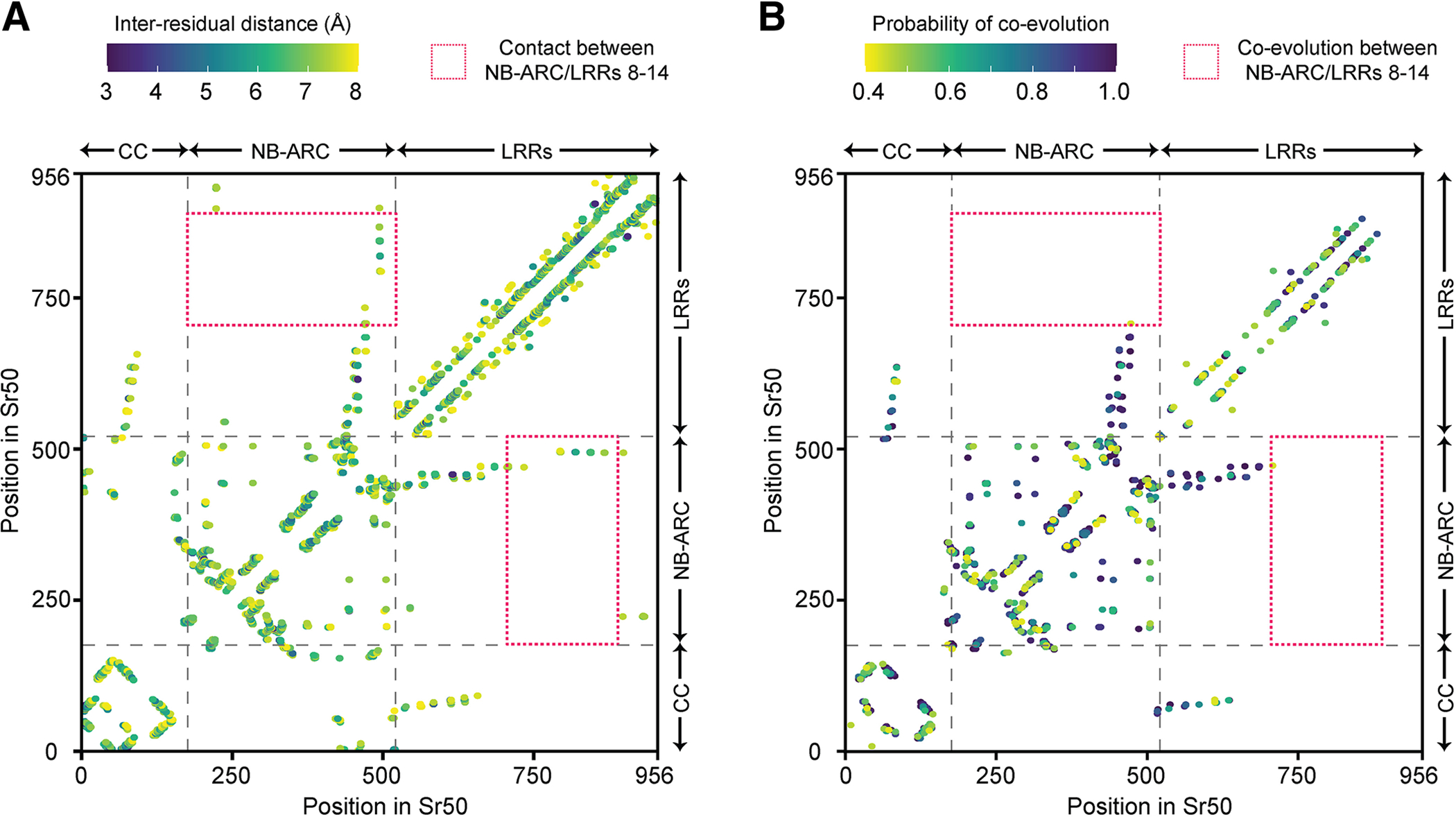
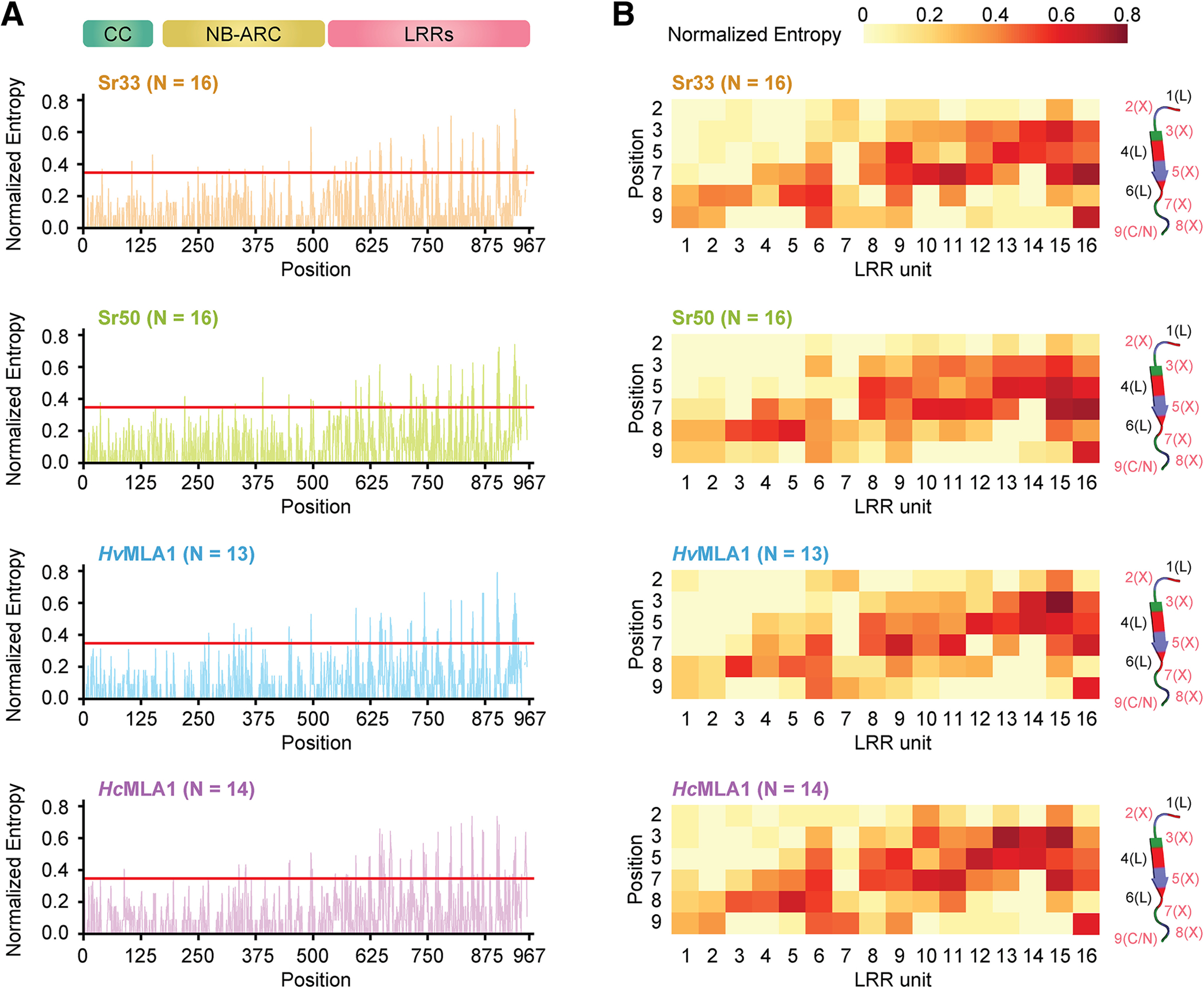
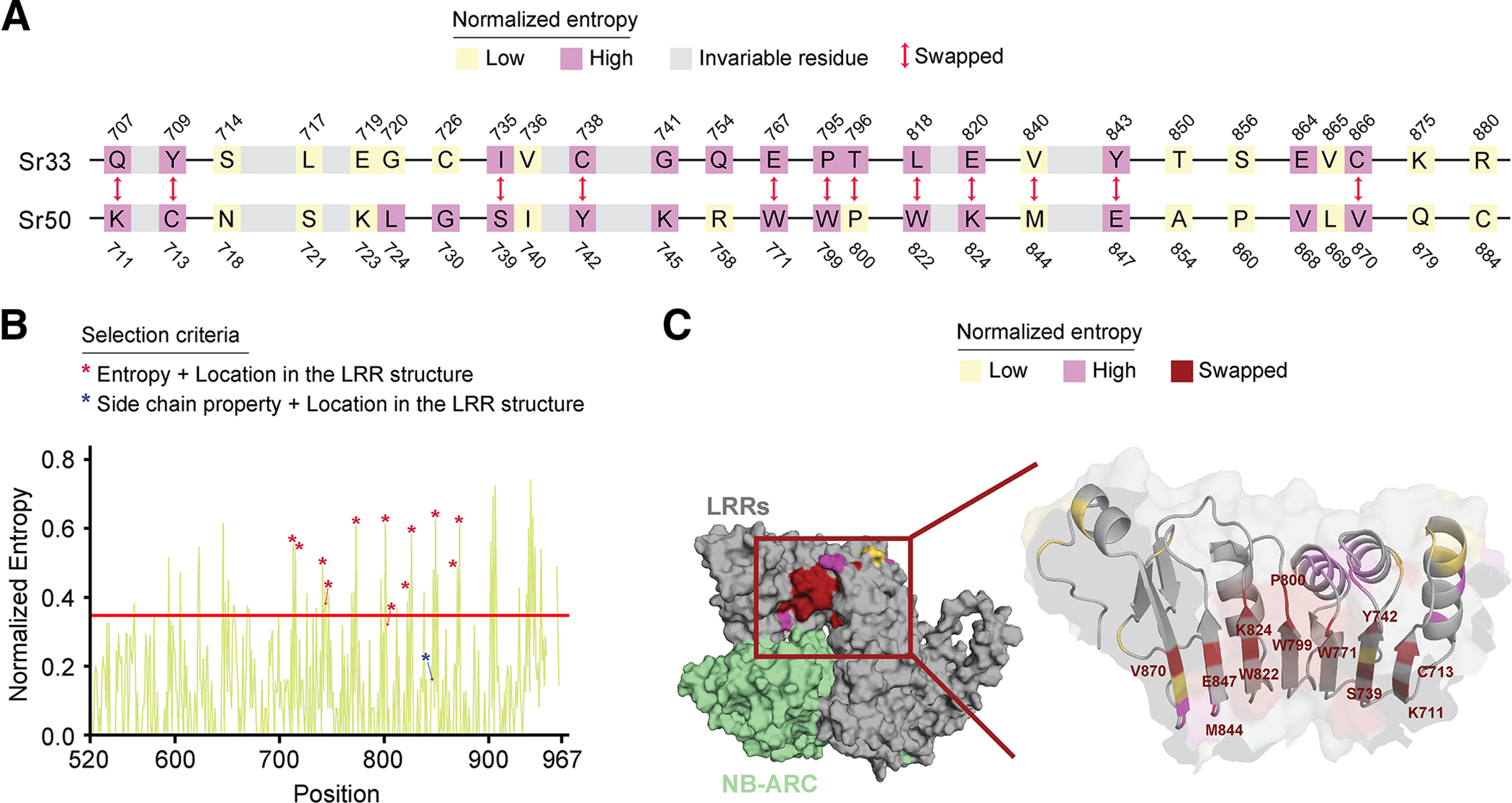
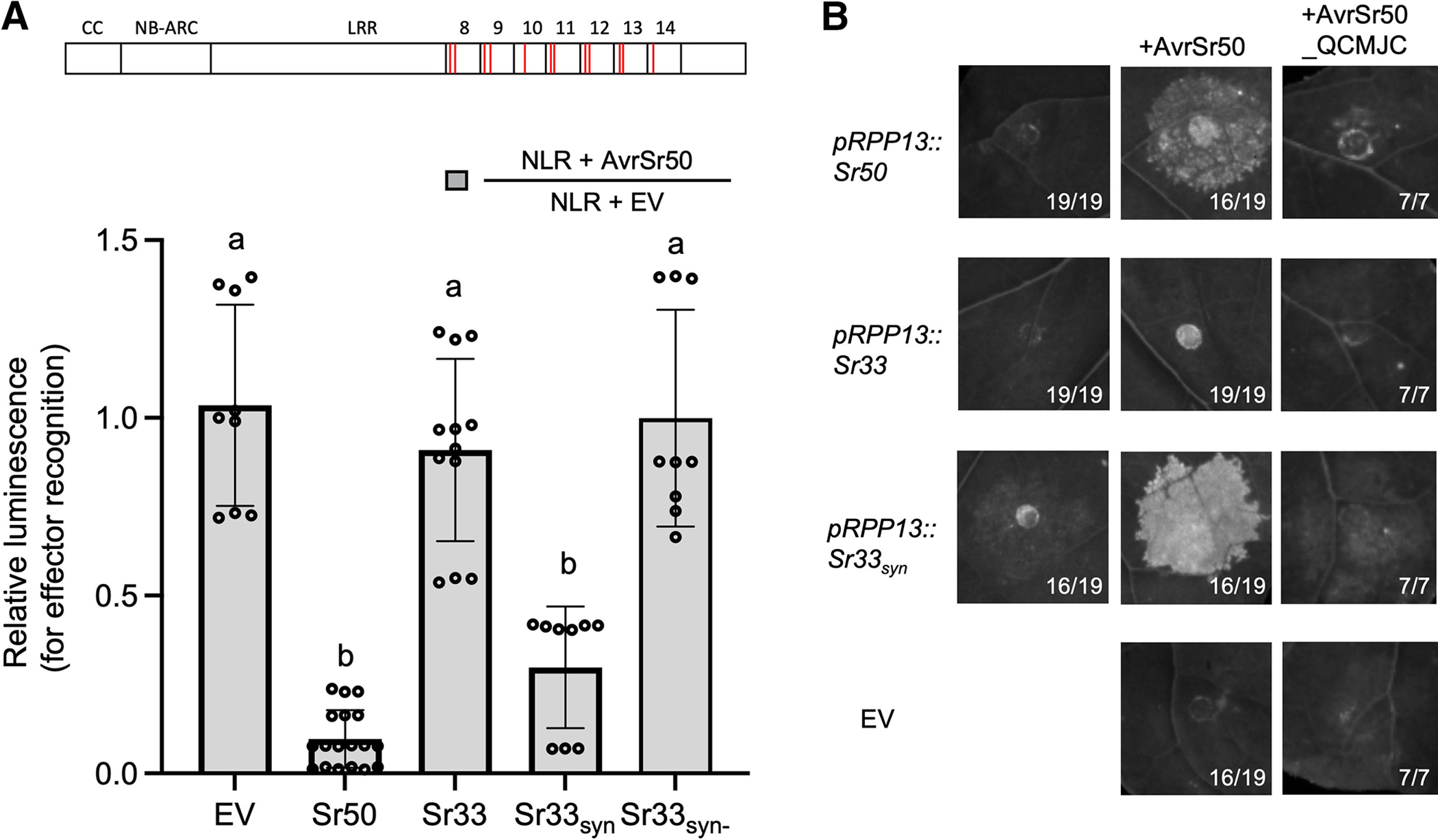
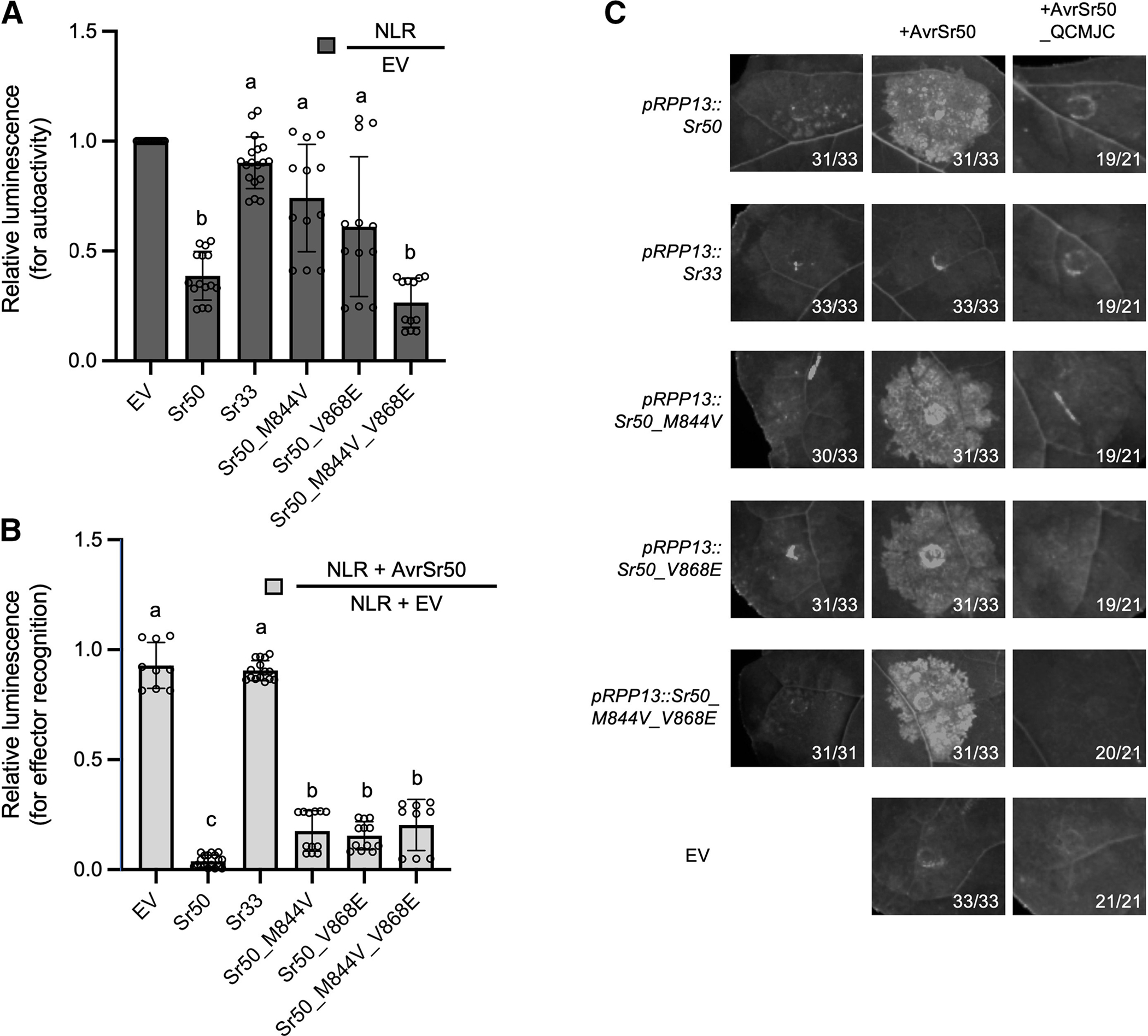
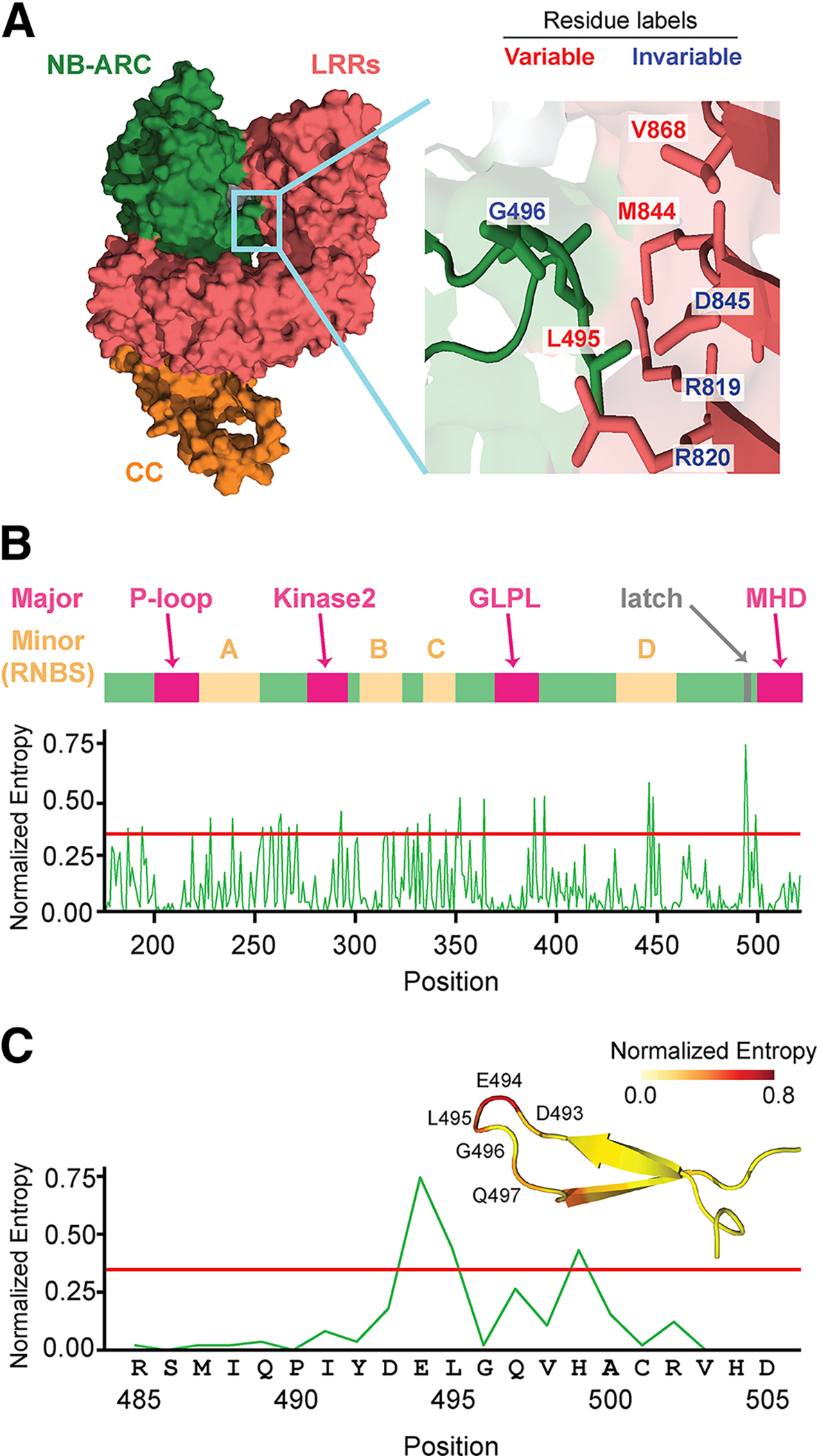
Many resistance genes deployed against pathogens in crops are intracellular nucleotide-binding (NB) leucine-rich repeat (LRR) receptors (NLRs). The ability to rationally engineer the specificity of NLRs will be crucial in the response to newly emerging crop diseases. Successful attempts to modify NLR recognition have been limited to untargeted approaches or depended on previously available structural information or knowledge of pathogen-effector targets. However, this information is not available for most NLR-effector pairs. Here, we demonstrate the precise prediction and subsequent transfer of residues involved in effector recognition between two closely related NLRs without their experimentally determined structure or detailed knowledge about their pathogen effector targets. By combining phylogenetics, allele diversity analysis, and structural modeling, we successfully predicted residues mediating interaction of Sr50 with its cognate effector AvrSr50 and transferred recognition specificity of Sr50 to the closely related NLR Sr33. We created synthetic versions of Sr33 that contain amino acids from Sr50, including Sr33, which gained the ability to recognize AvrSr50 with 12 amino-acid substitutions. Furthermore, we discovered that sites in the LRR domain needed to transfer recognition specificity to Sr33 also influence autoactivity in Sr50. Structural modeling suggests these residues interact with a part of the NB-ARC domain, which we named the NB-ARC latch, to possibly maintain the inactive state of the receptor. Our approach demonstrates rational modifications of NLRs, which could be useful to enhance existing elite crop germplasm. [Formula: see text] Copyright © 2023 The Author(s). This is an open access article distributed under the CC BY-NC-ND 4.0 International license.
许多用于对抗作物病原体的抗性基因是细胞内核苷酸结合(NB)亮氨酸重复(LRR)受体(NLRs)。合理设计 NLR 特异性的能力对于应对新出现的作物疾病至关重要。成功尝试修改 NLR 识别的方法仅限于无目标方法,或者依赖于以前可用的结构信息或病原体效应物靶标的知识。然而,对于大多数 NLR-效应物对,这些信息是不可用的。在这里,我们在没有 NLR 及其效应物靶标结构实验确定或详细知识的情况下,展示了两个密切相关的 NLR 之间涉及效应物识别的残基的精确预测和随后转移。通过结合系统发育学、等位基因多样性分析和结构建模,我们成功地预测了介导 Sr50 与其同源效应物 AvrSr50 相互作用的残基,并将 Sr50 的识别特异性转移到密切相关的 NLR Sr33 上。我们创建了 Sr33 的合成版本,其中包含来自 Sr50 的氨基酸,包括 Sr33,它能够通过 12 个氨基酸取代识别 AvrSr50。此外,我们发现需要将识别特异性转移到 Sr33 的 LRR 结构域中的位点也会影响 Sr50 的自动活性。结构建模表明,这些残基与 NB-ARC 结构域的一部分相互作用,我们将其命名为 NB-ARC 闩锁,以可能维持受体的非活性状态。我们的方法展示了 NLR 的合理修饰,这对于增强现有优良作物种质可能是有用的。