TC Jenkins Department of Biophysics, Johns Hopkins University, Baltimore, United States.
Laboratory of Cell and Developmental Signaling, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Frederick, United States.
Elife. 2023 Jul 12;12:e84881. doi: 10.7554/eLife.84881.
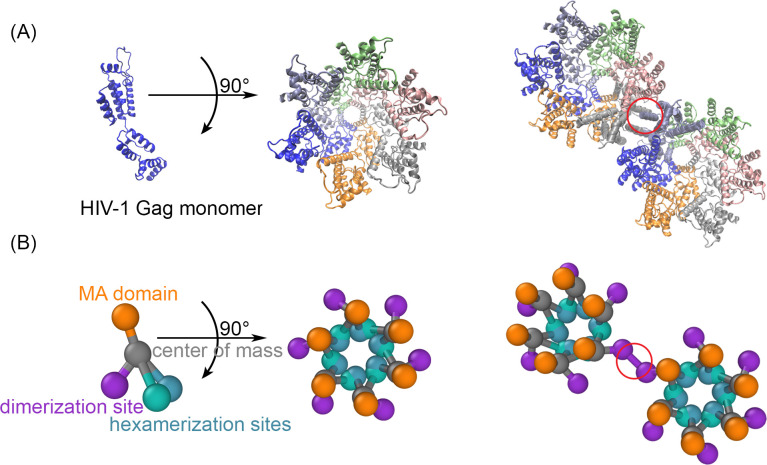
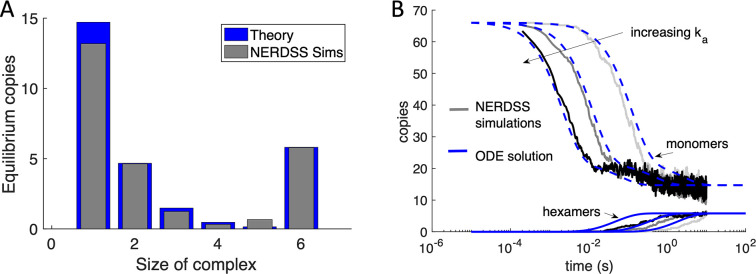
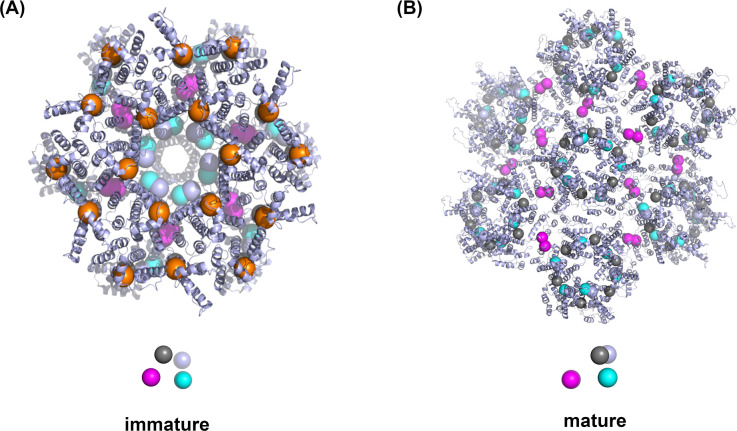
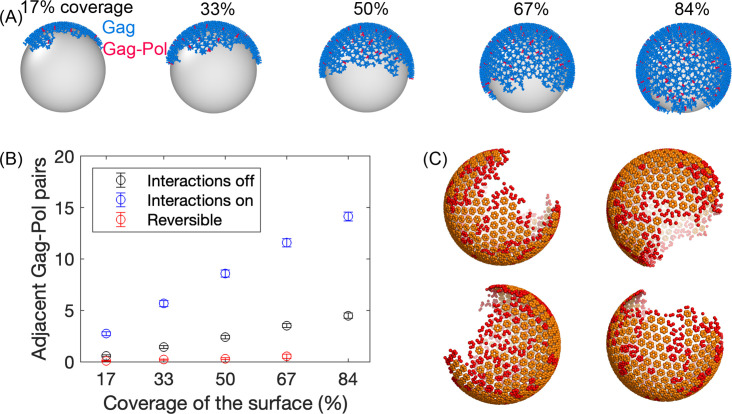
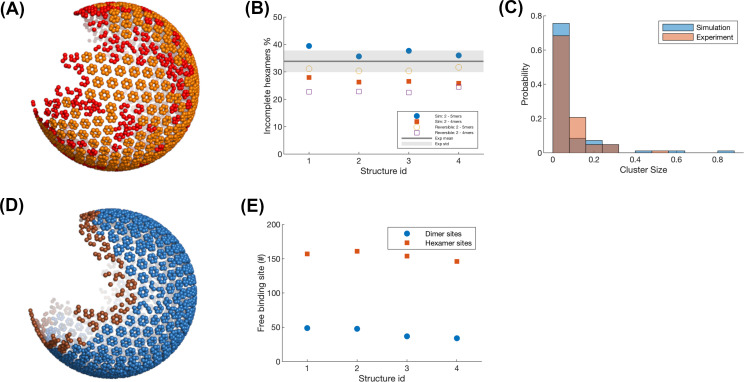
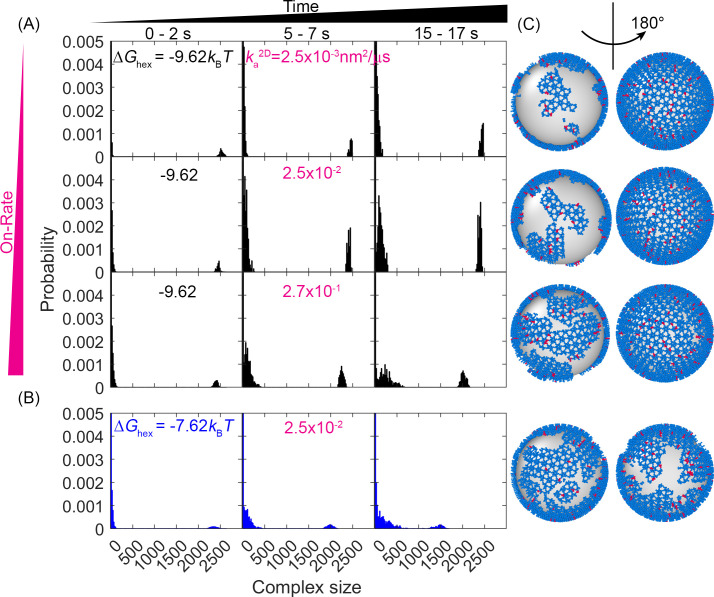
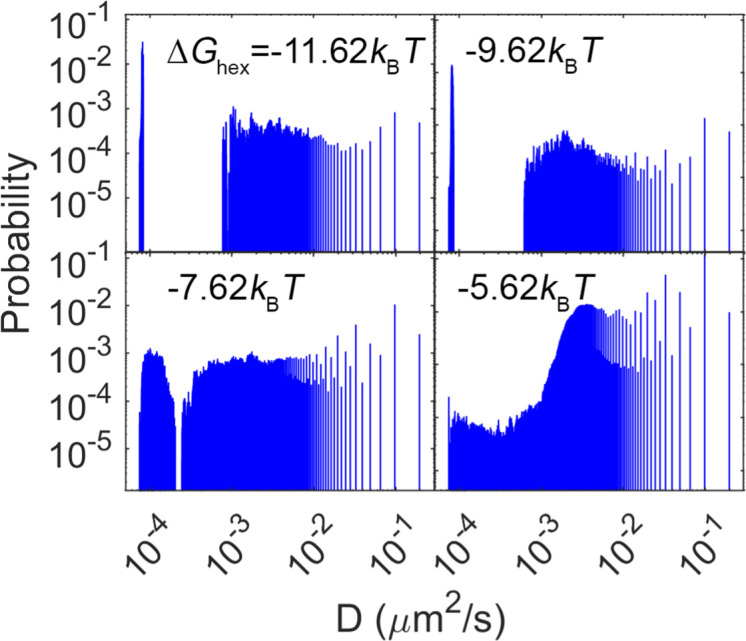
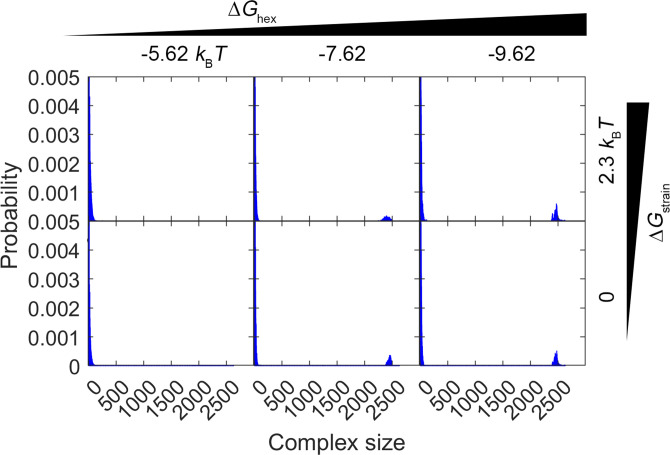
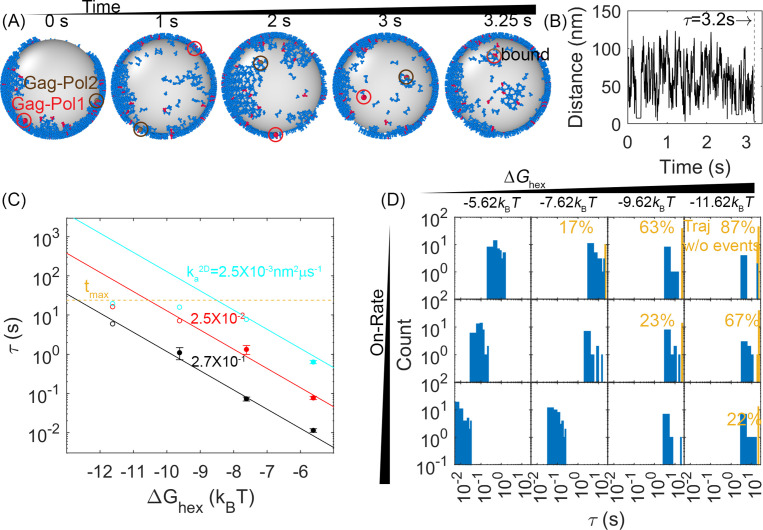
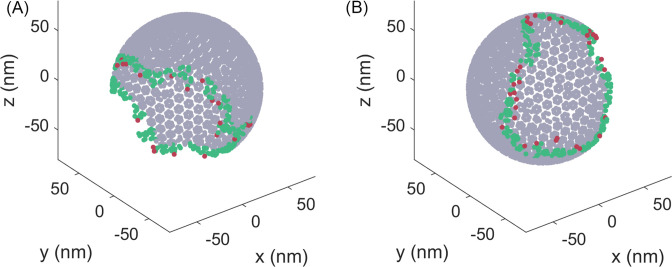
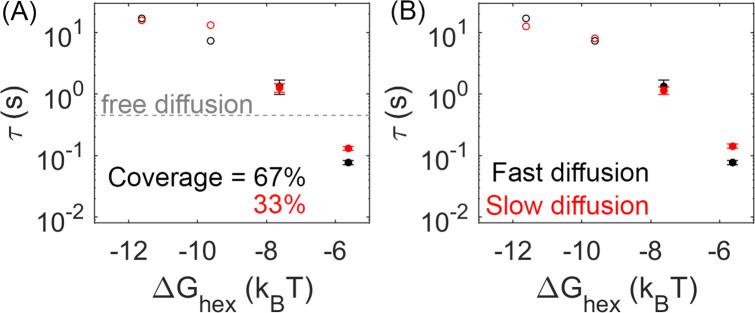
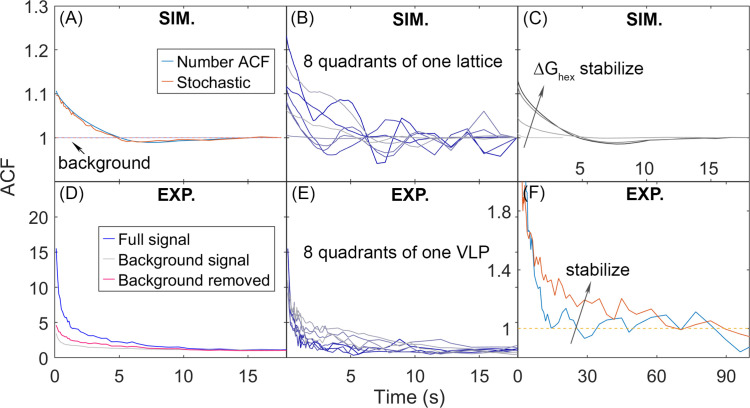
For HIV virions to become infectious, the immature lattice of Gag polyproteins attached to the virion membrane must be cleaved. Cleavage cannot initiate without the protease formed by the homo-dimerization of domains linked to Gag. However, only 5% of the Gag polyproteins, termed Gag-Pol, carry this protease domain, and they are embedded within the structured lattice. The mechanism of Gag-Pol dimerization is unknown. Here, we use spatial stochastic computer simulations of the immature Gag lattice as derived from experimental structures, showing that dynamics of the lattice on the membrane is unavoidable due to the missing 1/3 of the spherical protein coat. These dynamics allow for Gag-Pol molecules carrying the protease domains to detach and reattach at new places within the lattice. Surprisingly, dimerization timescales of minutes or less are achievable for realistic binding energies and rates despite retaining most of the large-scale lattice structure. We derive a formula allowing extrapolation of timescales as a function of interaction free energy and binding rate, thus predicting how additional stabilization of the lattice would impact dimerization times. We further show that during assembly, dimerization of Gag-Pol is highly likely and therefore must be actively suppressed to prevent early activation. By direct comparison to recent biochemical measurements within budded virions, we find that only moderately stable hexamer contacts (-12<∆<-8) retain both the dynamics and lattice structures that are consistent with experiment. These dynamics are likely essential for proper maturation, and our models quantify and predict lattice dynamics and protease dimerization timescales that define a key step in understanding formation of infectious viruses.
为了使 HIV 病毒成为感染性的,附着在病毒膜上的不成熟的 Gag 多聚蛋白晶格必须被切割。没有与 Gag 连接的结构域形成的同源二聚体蛋白酶的作用,切割就不能启动。然而,只有 5%的 Gag 多聚蛋白,称为 Gag-Pol,携带这种蛋白酶结构域,并且它们嵌入在结构晶格中。Gag-Pol 二聚化的机制尚不清楚。在这里,我们使用从实验结构中得出的不成熟 Gag 晶格的空间随机计算机模拟,表明由于缺少球形蛋白外壳的 1/3,膜上的晶格动力学是不可避免的。这些动力学允许携带蛋白酶结构域的 Gag-Pol 分子在晶格内的新位置上脱离和重新附着。令人惊讶的是,尽管保留了大部分大规模晶格结构,但在现实的结合能和速率下,实现分钟或更短的二聚化时间尺度是可行的。我们推导出一个公式,允许根据相互作用自由能和结合速率来推断时间尺度,从而预测晶格的额外稳定性如何影响二聚化时间。我们进一步表明,在装配过程中,Gag-Pol 的二聚化极有可能发生,因此必须积极抑制以防止过早激活。通过与最近在出芽病毒中进行的生化测量的直接比较,我们发现只有中等稳定的六聚体接触(-12<∆<-8)保留了与实验一致的动力学和晶格结构。这些动力学可能对适当的成熟至关重要,我们的模型量化并预测了定义理解传染性病毒形成的关键步骤的晶格动力学和蛋白酶二聚化时间尺度。