Berndsen Christopher E, Storm Amanda R, Sardelli Angelina M, Hossain Sheikh R, Clermont Kristen R, McFather Luke M, Connor Mafe A, Monroe Jonathan D
Department of Chemistry and Biochemistry, James Madison University, Harrisonburg, VA 22807.
Department of Biology, Western Carolina University, Cullowhee, NC 28723.
bioRxiv. 2024 Aug 7:2024.08.07.607052. doi: 10.1101/2024.08.07.607052.
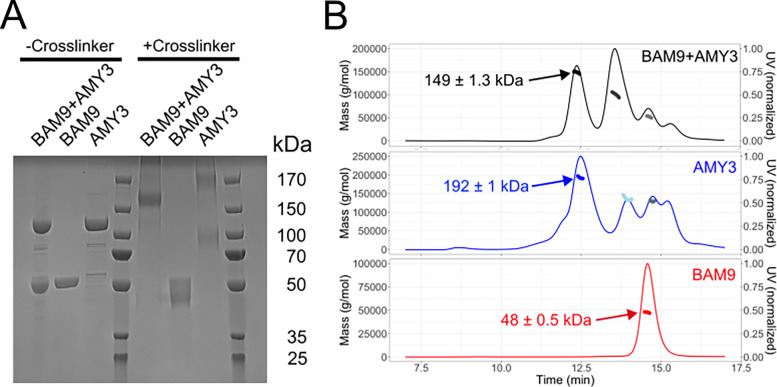
Starch accumulation in plant tissues provides an important carbon source at night and for regrowth after periods of dormancy and in times of stress. Both ɑ- and β-amylases (AMYs and BAMs, respectively) catalyze starch hydrolysis, but their functional roles are unclear. Moreover, the presence of catalytically inactive amylases that show starch excess phenotypes when deleted presents an interesting series of questions on how starch degradation is regulated. Plants lacking one of these catalytically inactive β-amylases, BAM9, were shown to have enhanced starch accumulation when combined with mutations in BAM1 and BAM3, the primary starch degrading BAMs in response to stress and at night, respectively. Importantly, BAM9 has been reported to be transcriptionally induced by stress through activation of SnRK1. Using yeast two-hybrid experiments, we identified the plastid-localized AMY3 as a potential interaction partner for BAM9. We found that BAM9 interacted with AMY3 and that BAM9 enhances AMY3 activity 3-fold. Modeling of the AMY3-BAM9 complex revealed a previously undescribed N-terminal structural feature in AMY3 that we call the alpha-alpha hairpin that could serve as a potential interaction site. Additionally, AMY3 lacking the alpha-alpha hairpin is unaffected by BAM9. Structural analysis of AMY3 showed that it can form a homodimer in solution and that BAM9 appears to replace one of the AMY3 monomers to form a heterodimer. Collectively these data suggest that BAM9 is a pseudoamylase that activates AMY3 in response to cellular stress, possibly facilitating starch degradation to provide an additional energy source for stress recovery.
植物组织中的淀粉积累为夜间以及休眠期后和应激时的再生长提供了重要的碳源。α-淀粉酶和β-淀粉酶(分别为AMYs和BAMs)均催化淀粉水解,但其功能作用尚不清楚。此外,存在一些催化无活性的淀粉酶,当它们缺失时会表现出淀粉过量的表型,这就淀粉降解是如何调控的提出了一系列有趣的问题。研究表明,缺乏其中一种催化无活性的β-淀粉酶BAM9的植物,与BAM1和BAM3的突变体结合时,淀粉积累会增强,BAM1和BAM3分别是应激时和夜间主要的淀粉降解BAMs。重要的是,据报道BAM9通过SnRK1的激活而受到应激的转录诱导。通过酵母双杂交实验,我们确定了定位于质体的AMY3是BAM9的潜在相互作用伙伴。我们发现BAM9与AMY3相互作用,并且BAM9使AMY3的活性提高了3倍。AMY3 - BAM9复合物的模型揭示了AMY3中一个以前未描述的N端结构特征,我们称之为α-α发夹结构,它可能作为一个潜在的相互作用位点。此外,缺乏α-α发夹结构的AMY3不受BAM9的影响。AMY3的结构分析表明,它在溶液中可以形成同二聚体,并且BAM9似乎取代了AMY3的一个单体以形成异二聚体。这些数据共同表明,BAM9是一种假淀粉酶,它在细胞应激时激活AMY3,可能促进淀粉降解以提供额外的能量来源用于应激恢复。