Svitkina T M, Verkhovsky A B, McQuade K M, Borisy G G
Laboratory of Molecular Biology, University of Wisconsin, Madison, Wisconsin 53706, USA.
J Cell Biol. 1997 Oct 20;139(2):397-415. doi: 10.1083/jcb.139.2.397.
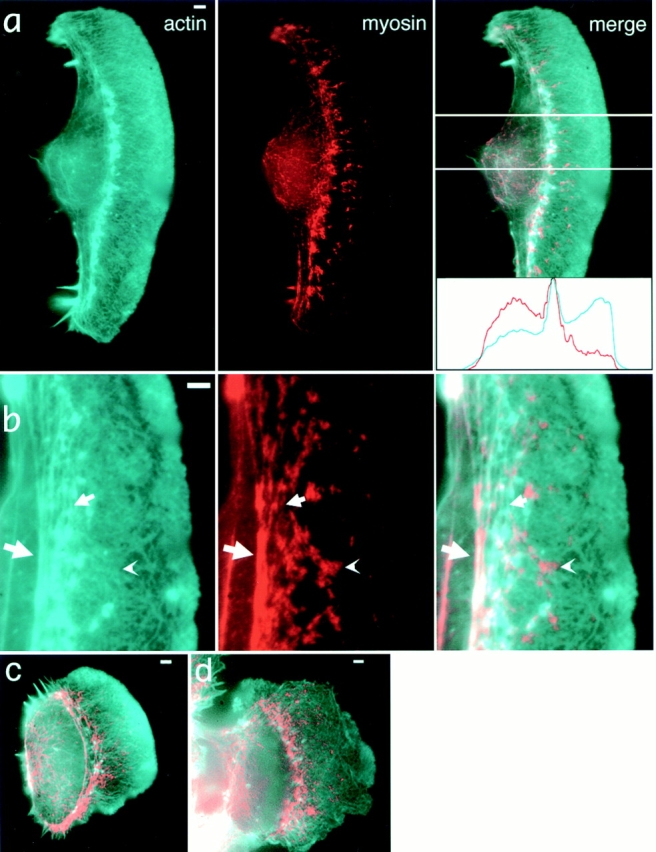
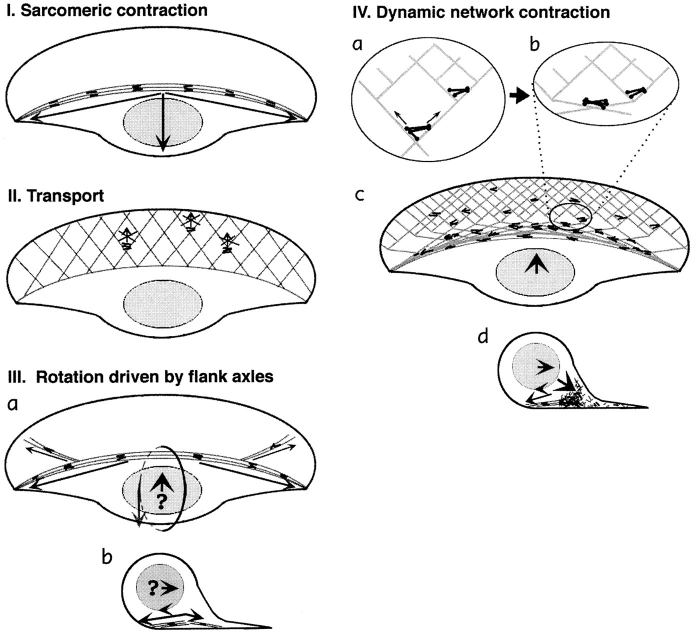
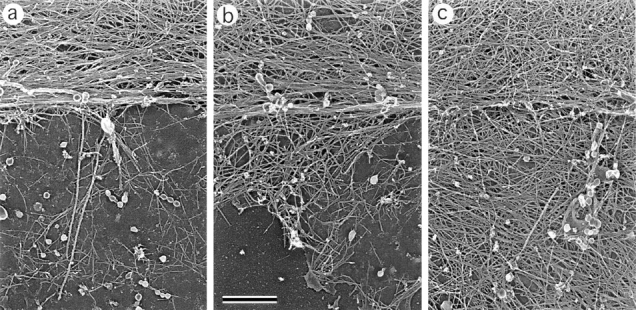
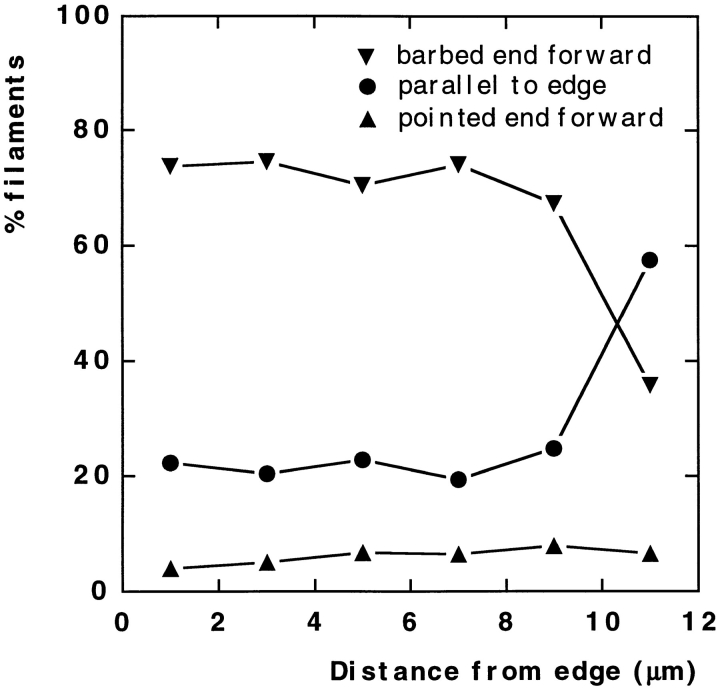
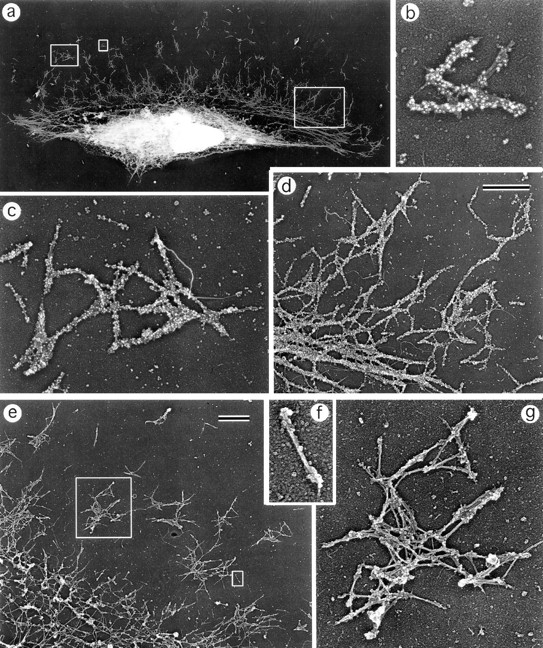
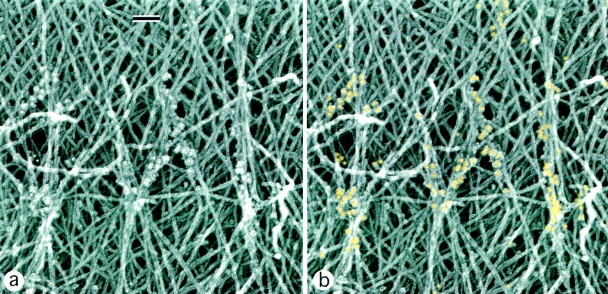
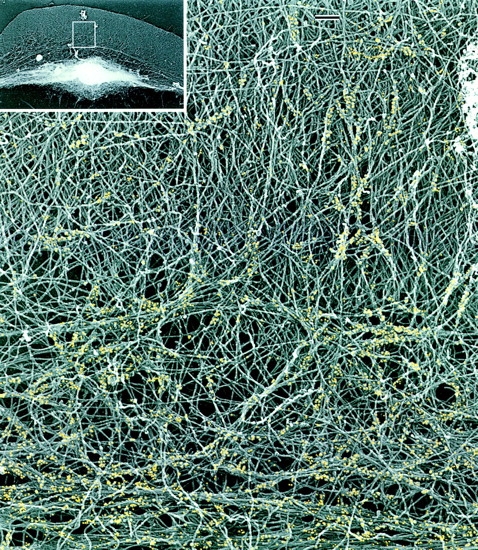
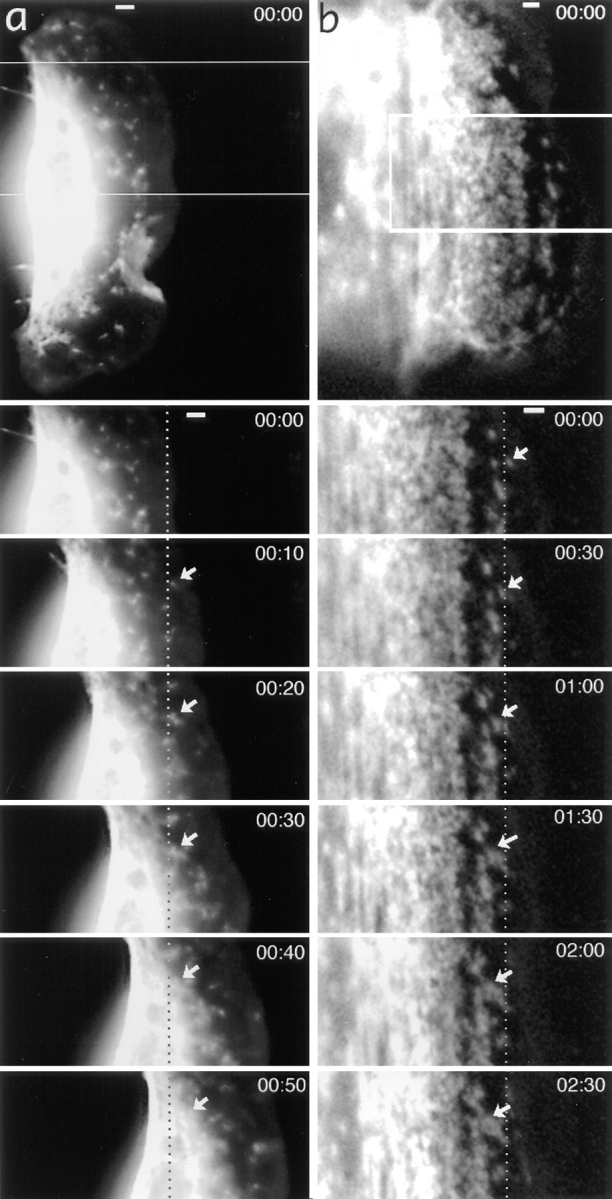
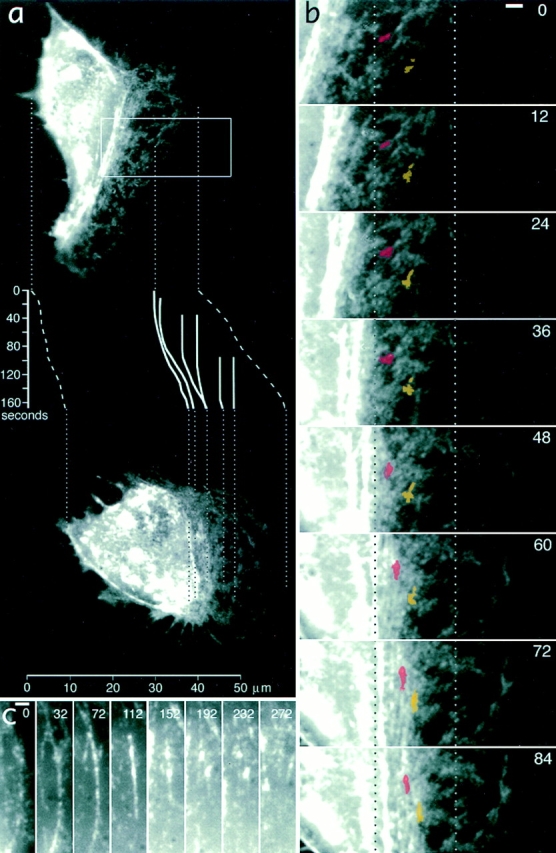
While the protrusive event of cell locomotion is thought to be driven by actin polymerization, the mechanism of forward translocation of the cell body is unclear. To elucidate the mechanism of cell body translocation, we analyzed the supramolecular organization of the actin-myosin II system and the dynamics of myosin II in fish epidermal keratocytes. In lamellipodia, long actin filaments formed dense networks with numerous free ends in a brushlike manner near the leading edge. Shorter actin filaments often formed T junctions with longer filaments in the brushlike area, suggesting that new filaments could be nucleated at sides of preexisting filaments or linked to them immediately after nucleation. The polarity of actin filaments was almost uniform, with barbed ends forward throughout most of the lamellipodia but mixed in arc-shaped filament bundles at the lamellipodial/cell body boundary. Myosin II formed discrete clusters of bipolar minifilaments in lamellipodia that increased in size and density towards the cell body boundary and colocalized with actin in boundary bundles. Time-lapse observation demonstrated that myosin clusters appeared in the lamellipodia and remained stationary with respect to the substratum in locomoting cells, but they exhibited retrograde flow in cells tethered in epithelioid colonies. Consequently, both in locomoting and stationary cells, myosin clusters approached the cell body boundary, where they became compressed and aligned, resulting in the formation of boundary bundles. In locomoting cells, the compression was associated with forward displacement of myosin features. These data are not consistent with either sarcomeric or polarized transport mechanisms of cell body translocation. We propose that the forward translocation of the cell body and retrograde flow in the lamellipodia are both driven by contraction of an actin-myosin network in the lamellipodial/cell body transition zone.
虽然细胞运动的突出事件被认为是由肌动蛋白聚合驱动的,但细胞体向前移位的机制尚不清楚。为了阐明细胞体移位的机制,我们分析了鱼类表皮角质形成细胞中肌动蛋白-肌球蛋白II系统的超分子组织和肌球蛋白II的动力学。在片状伪足中,长肌动蛋白丝在靠近前沿的位置以刷状方式形成密集网络,有许多自由末端。较短的肌动蛋白丝在刷状区域常与较长的肌动蛋白丝形成T形连接,这表明新的肌动蛋白丝可能在现有肌动蛋白丝的侧面成核,或者在成核后立即与它们相连。肌动蛋白丝的极性几乎是均匀的,在大多数片状伪足中,带刺末端向前,但在片状伪足/细胞体边界处的弧形丝束中则是混合的。肌球蛋白II在片状伪足中形成双极微丝的离散簇,其大小和密度朝着细胞体边界增加,并与边界束中的肌动蛋白共定位。延时观察表明,肌球蛋白簇出现在片状伪足中,在运动细胞中相对于基质保持静止,但在附着于上皮样菌落的细胞中它们表现出逆行流动。因此,在运动细胞和静止细胞中,肌球蛋白簇都接近细胞体边界,在那里它们被压缩并排列,导致边界束的形成。在运动细胞中,压缩与肌球蛋白特征的向前位移有关。这些数据与细胞体移位的肌节或极化运输机制均不一致。我们提出,细胞体的向前移位和片状伪足中的逆行流动都是由片状伪足/细胞体过渡区的肌动蛋白-肌球蛋白网络收缩驱动的。