Poolman B, Molenaar D, Smid E J, Ubbink T, Abee T, Renault P P, Konings W N
Department of Microbiology, University of Groningen, Haren, The Netherlands.
J Bacteriol. 1991 Oct;173(19):6030-7. doi: 10.1128/jb.173.19.6030-6037.1991.
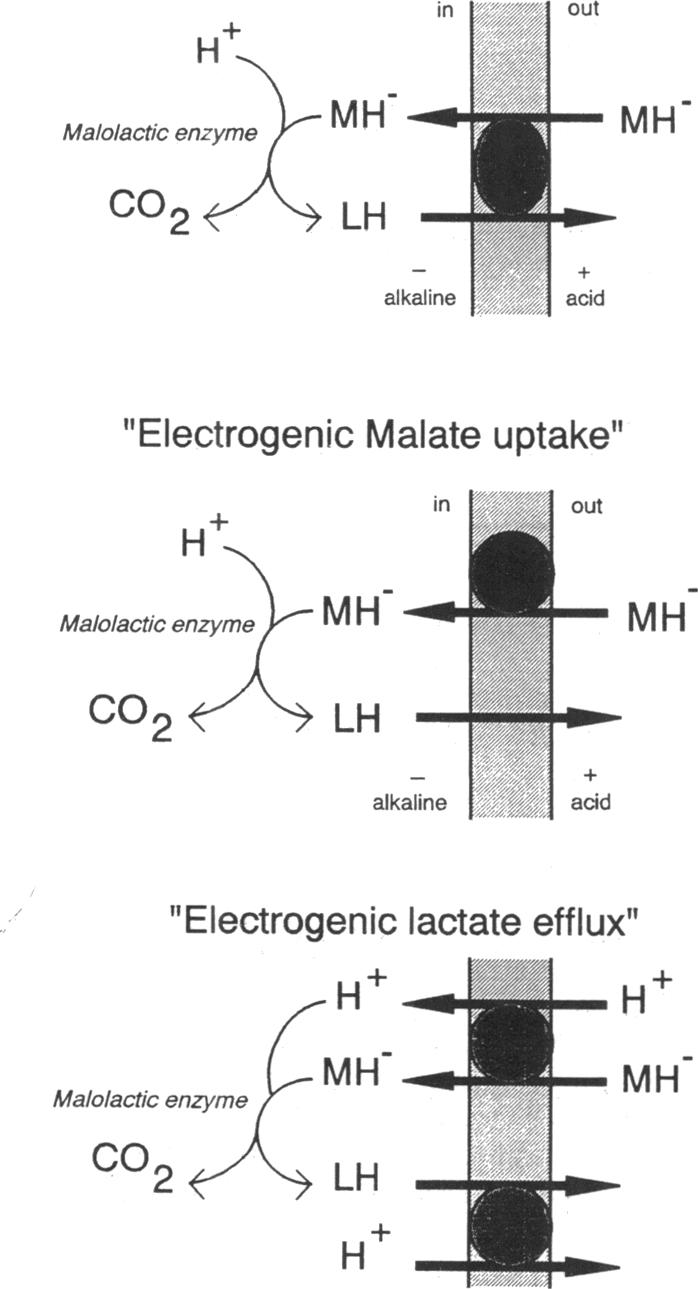
The mechanism of metabolic energy production by malolactic fermentation in Lactococcus lactis has been investigated. In the presence of L-malate, a proton motive force composed of a membrane potential and pH gradient is generated which has about the same magnitude as the proton motive force generated by the metabolism of a glycolytic substrate. Malolactic fermentation results in the synthesis of ATP which is inhibited by the ionophore nigericin and the F0F1-ATPase inhibitor N,N-dicyclohexylcarbodiimide. Since substrate-level phosphorylation does not occur during malolactic fermentation, the generation of metabolic energy must originate from the uptake of L-malate and/or excretion of L-lactate. The initiation of malolactic fermentation is stimulated by the presence of L-lactate intracellularly, suggesting that L-malate is exchanged for L-lactate. Direct evidence for heterologous L-malate/L-lactate (and homologous L-malate/L-malate) antiport has been obtained with membrane vesicles of an L. lactis mutant deficient in malolactic enzyme. In membrane vesicles fused with liposomes, L-malate efflux and L-malate/L-lactate antiport are stimulated by a membrane potential (inside negative), indicating that net negative charge is moved to the outside in the efflux and antiport reaction. In membrane vesicles fused with liposomes in which cytochrome c oxidase was incorporated as a proton motive force-generating mechanism, transport of L-malate can be driven by a pH gradient alone, i.e., in the absence of L-lactate as countersubstrate. A membrane potential (inside negative) inhibits uptake of L-malate, indicating that L-malate is transported an an electronegative monoanionic species (or dianionic species together with a proton). The experiments described suggest that the generation of metabolic energy during malolactic fermentation arises from electrogenic malate/lactate antiport and electrogenic malate uptake (in combination with outward diffusion of lactic acid), together with proton consumption as result of decarboxylation of L-malate. The net energy gain would be equivalent to one proton translocated form the inside to the outside per L-malate metabolized.
对乳酸乳球菌中苹果酸-乳酸发酵产生代谢能量的机制进行了研究。在L-苹果酸存在的情况下,会产生由膜电位和pH梯度组成的质子动力,其大小与糖酵解底物代谢产生的质子动力大致相同。苹果酸-乳酸发酵导致ATP的合成,该合成受到离子载体尼日利亚菌素和F0F1-ATP酶抑制剂N,N-二环己基碳二亚胺的抑制。由于在苹果酸-乳酸发酵过程中不发生底物水平磷酸化,代谢能量的产生必定源于L-苹果酸的摄取和/或L-乳酸的排泄。细胞内L-乳酸的存在会刺激苹果酸-乳酸发酵的启动,这表明L-苹果酸与L-乳酸发生了交换。利用缺乏苹果酸-乳酸酶的乳酸乳球菌突变体的膜囊泡,已获得了异源L-苹果酸/L-乳酸(以及同源L-苹果酸/L-苹果酸)反向转运的直接证据。在与脂质体融合的膜囊泡中,膜电位(内部为负)会刺激L-苹果酸外流和L-苹果酸/L-乳酸反向转运,这表明在流出和反向转运反应中净负电荷转移到了外部。在与掺入细胞色素c氧化酶作为质子动力产生机制的脂质体融合的膜囊泡中,L-苹果酸的转运可以仅由pH梯度驱动,即在没有L-乳酸作为反向底物的情况下。膜电位(内部为负)会抑制L-苹果酸的摄取,这表明L-苹果酸是以带负电的单阴离子形式(或与质子一起的双阴离子形式)进行转运的。所述实验表明,苹果酸-乳酸发酵过程中代谢能量的产生源于电生性苹果酸/乳酸反向转运和电生性苹果酸摄取(与乳酸的向外扩散相结合),以及L-苹果酸脱羧导致的质子消耗。每代谢一分子L-苹果酸,净能量增益相当于一个质子从内部转运到外部。