Institute of Virology, Hannover Medical School, Hannover, Germany.
PLoS Pathog. 2010 Jul 8;6(7):e1000991. doi: 10.1371/journal.ppat.1000991.
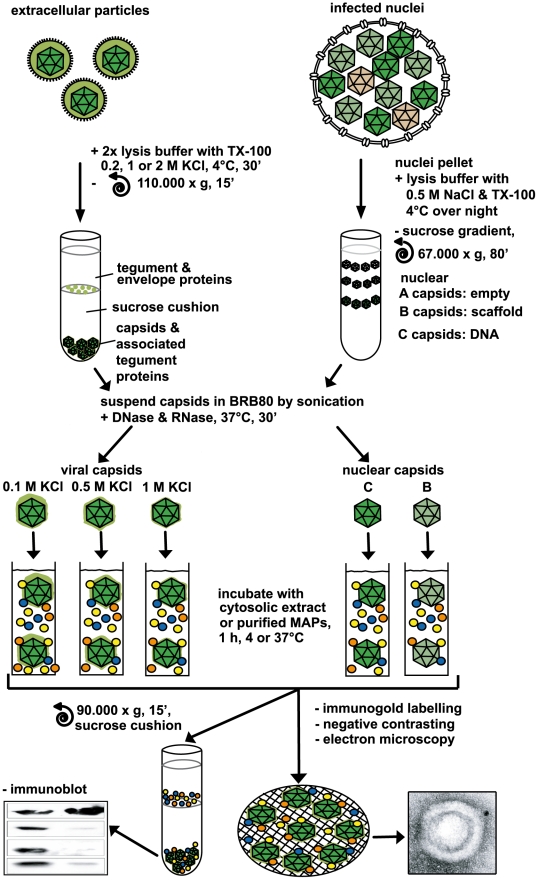
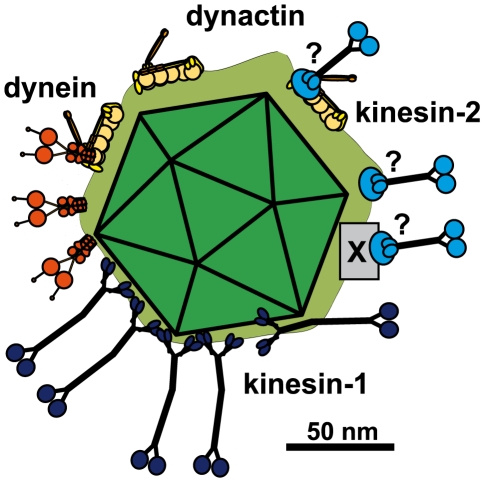
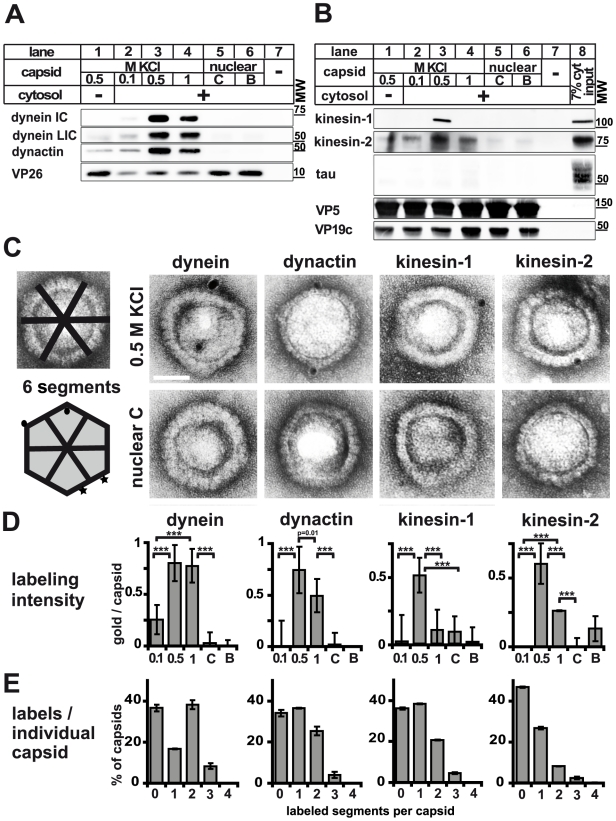
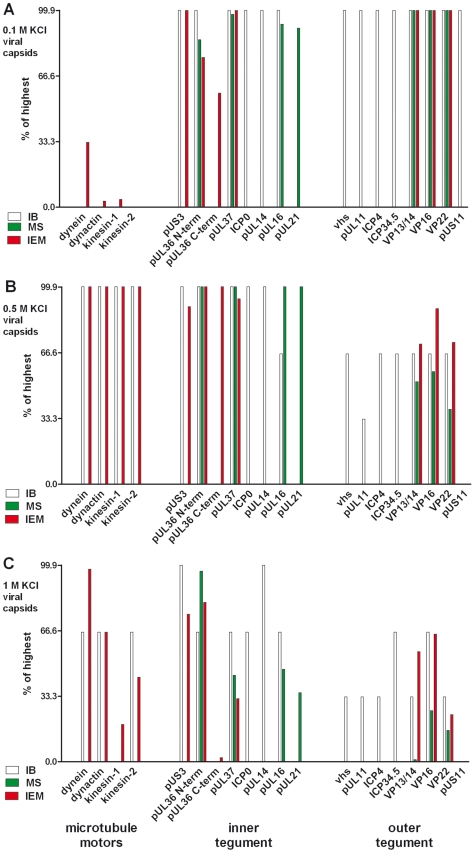
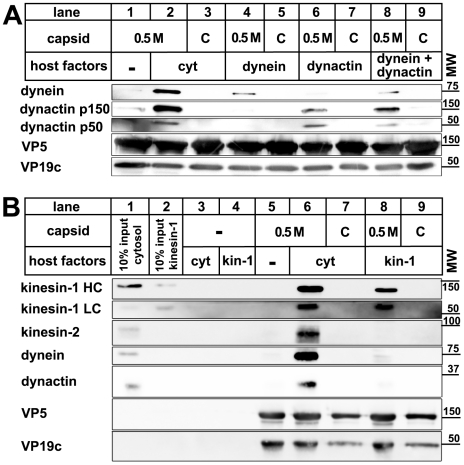
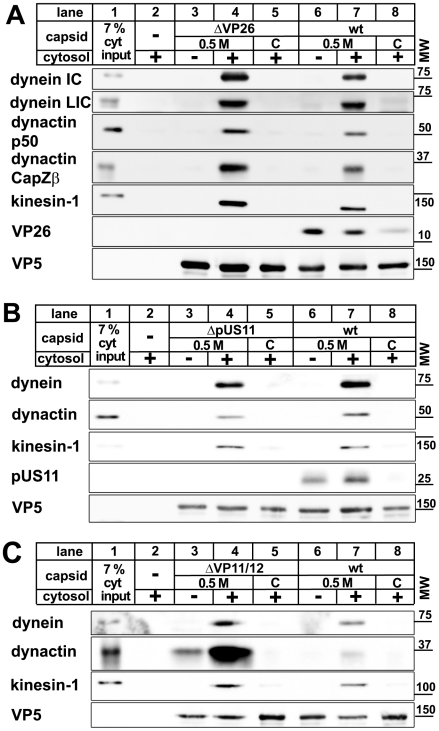
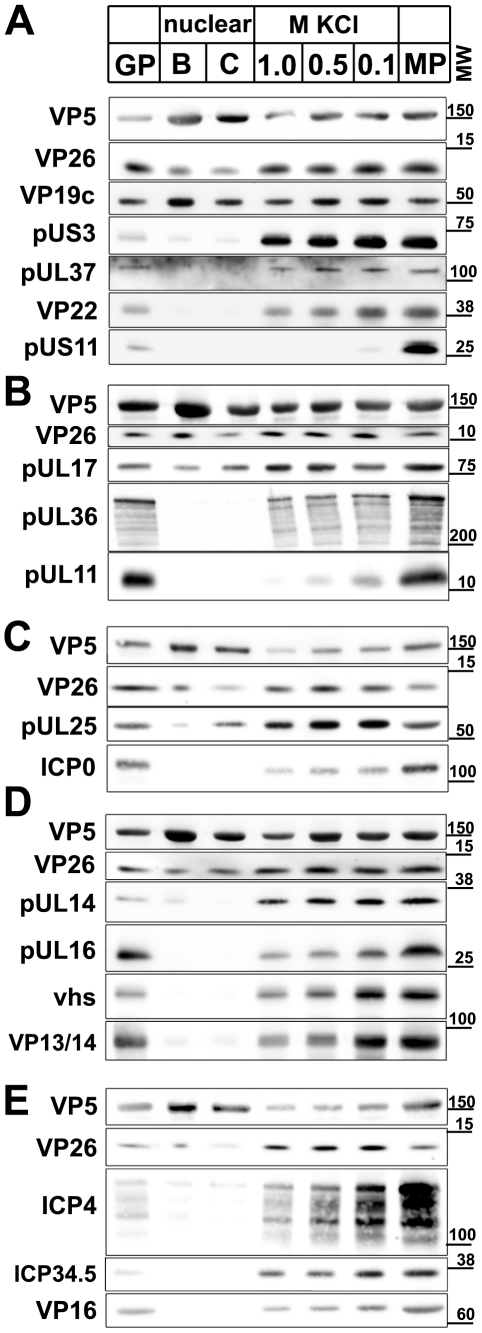
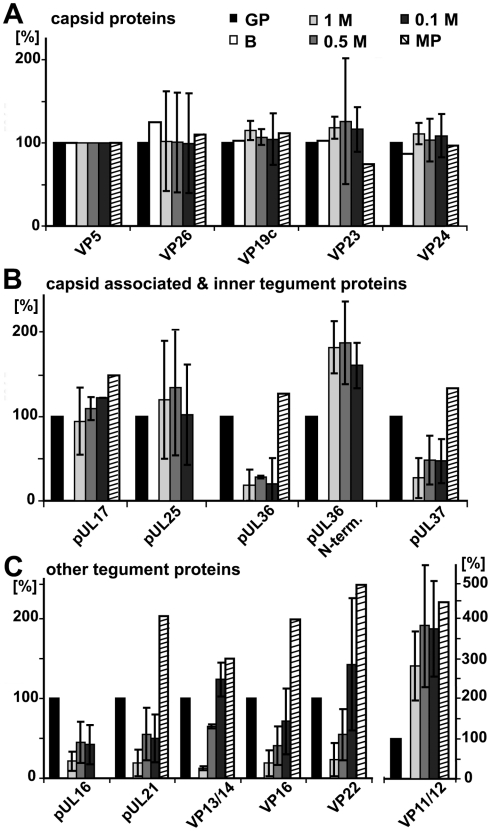
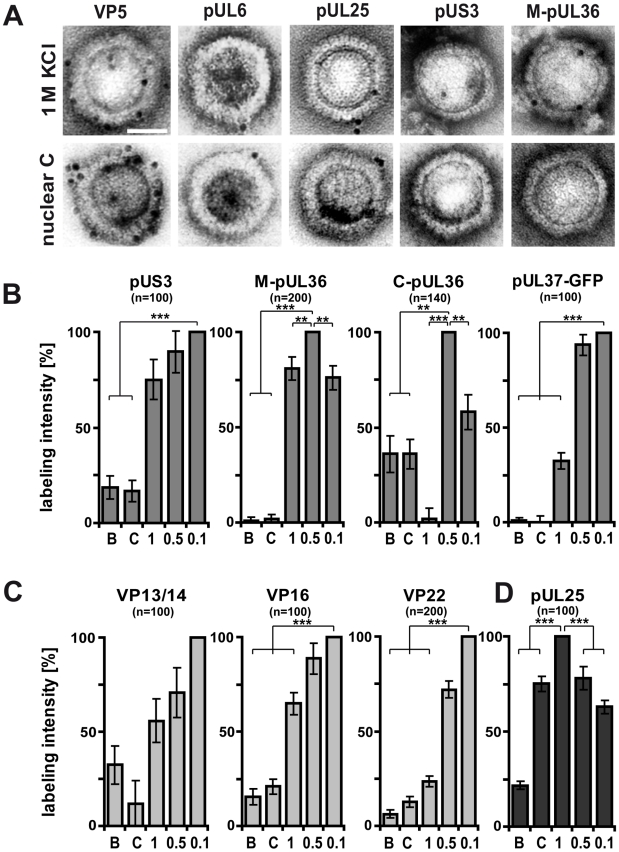
Many viruses depend on host microtubule motors to reach their destined intracellular location. Viral particles of neurotropic alphaherpesviruses such as herpes simplex virus 1 (HSV1) show bidirectional transport towards the cell center as well as the periphery, indicating that they utilize microtubule motors of opposing directionality. To understand the mechanisms of specific motor recruitment, it is necessary to characterize the molecular composition of such motile viral structures. We have generated HSV1 capsids with different surface features without impairing their overall architecture, and show that in a mammalian cell-free system the microtubule motors dynein and kinesin-1 and the dynein cofactor dynactin could interact directly with capsids independent of other host factors. The capsid composition and surface was analyzed with respect to 23 structural proteins that are potentially exposed to the cytosol during virus assembly or cell entry. Many of these proteins belong to the tegument, the hallmark of all herpesviruses located between the capsid and the viral envelope. Using immunoblots, quantitative mass spectrometry and quantitative immunoelectron microscopy, we show that capsids exposing inner tegument proteins such as pUS3, pUL36, pUL37, ICP0, pUL14, pUL16, and pUL21 recruited dynein, dynactin, kinesin-1 and kinesin-2. In contrast, neither untegumented capsids exposing VP5, VP26, pUL17 and pUL25 nor capsids covered by outer tegument proteins such as vhs, pUL11, ICP4, ICP34.5, VP11/12, VP13/14, VP16, VP22 or pUS11 bound microtubule motors. Our data suggest that HSV1 uses different structural features of the inner tegument to recruit dynein or kinesin-1. Individual capsids simultaneously accommodated motors of opposing directionality as well as several copies of the same motor. Thus, these associated motors either engage in a tug-of-war or their activities are coordinately regulated to achieve net transport either to the nucleus during cell entry or to cytoplasmic membranes for envelopment during assembly.
许多病毒依赖于宿主微管马达来到达其内在的细胞内位置。神经嗜性α疱疹病毒(如单纯疱疹病毒 1[HSV1])的病毒颗粒显示出朝向细胞中心和外周的双向运输,表明它们利用了相反方向的微管马达。为了理解特定马达募集的机制,有必要对这种运动病毒结构的分子组成进行表征。我们生成了具有不同表面特征的 HSV1 衣壳,而不会损害其整体结构,并表明在哺乳动物无细胞系统中,微管马达动力蛋白和驱动蛋白-1以及动力蛋白共因子 dynactin 可以直接与衣壳相互作用,而无需其他宿主因子。对衣壳组成和表面进行了分析,涉及 23 种结构蛋白,这些蛋白在病毒组装或细胞进入期间可能暴露于细胞质中。其中许多蛋白属于所有疱疹病毒的标志性结构——位于衣壳和病毒包膜之间的被膜。使用免疫印迹、定量质谱和定量免疫电子显微镜,我们表明,暴露于内被膜蛋白(如 pUS3、pUL36、pUL37、ICP0、pUL14、pUL16 和 pUL21)的衣壳招募了动力蛋白、dynactin、驱动蛋白-1 和驱动蛋白-2。相比之下,既未被膜暴露的衣壳(暴露 VP5、VP26、pUL17 和 pUL25),也未被包膜蛋白(如 vhs、pUL11、ICP4、ICP34.5、VP11/12、VP13/14、VP16、VP22 或 pUS11)覆盖的衣壳不结合微管马达。我们的数据表明,HSV1 利用内被膜的不同结构特征来募集动力蛋白或驱动蛋白-1。单个衣壳同时容纳了相反方向的马达以及相同马达的多个副本。因此,这些相关的马达要么参与拔河比赛,要么它们的活动被协调调节,以实现进入细胞时向核的净运输,或在组装时向细胞质膜的包膜运输。