Department of Genetics, Evolution and Environment and UCL Cancer Institute, University College London, London WC1E 6BT, UK.
Genome Biol. 2011 Aug 22;12(8):R82. doi: 10.1186/gb-2011-12-8-r82.
The generation of mature mRNAs involves interconnected processes, including transcription by RNA polymerase II (Pol II), modification of histones, and processing of pre-mRNAs through capping, intron splicing, and polyadenylation. These processes are thought to be integrated, both spatially and temporally, but it is unclear how these connections manifest at a global level with respect to chromatin patterns and transcription kinetics. We sought to clarify the relationships between chromatin, transcription and splicing using multiple genome-wide approaches in fission yeast.
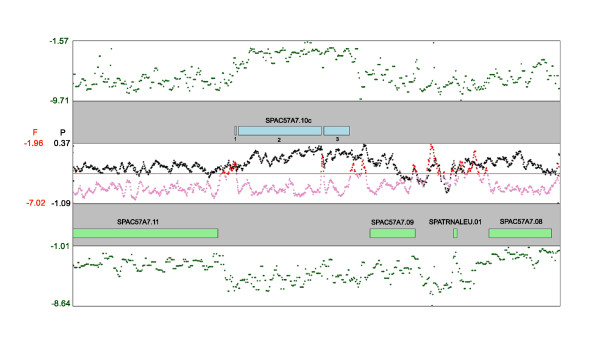
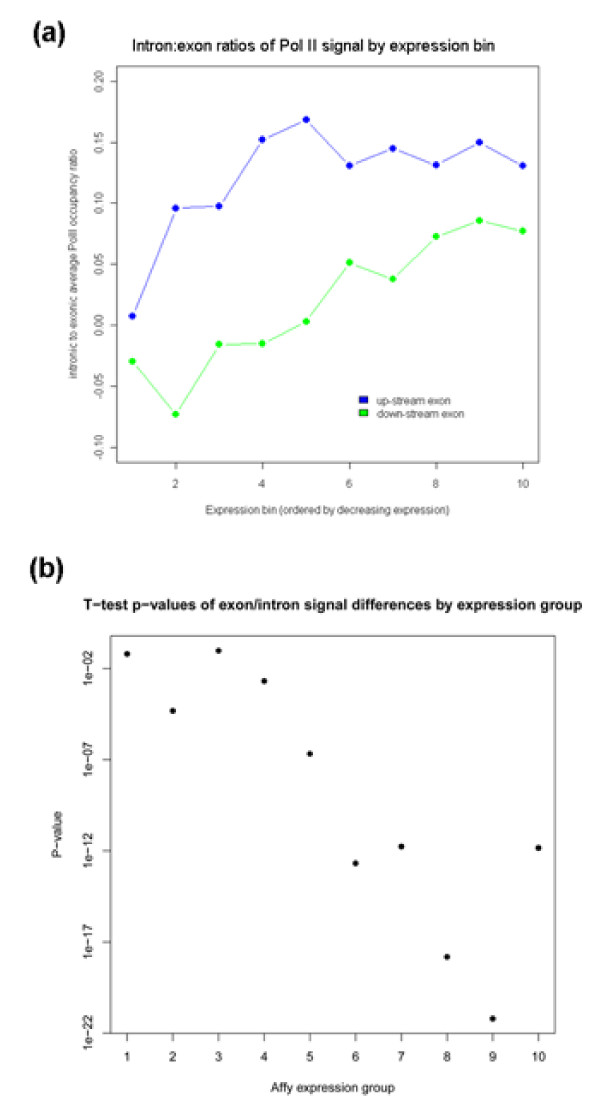
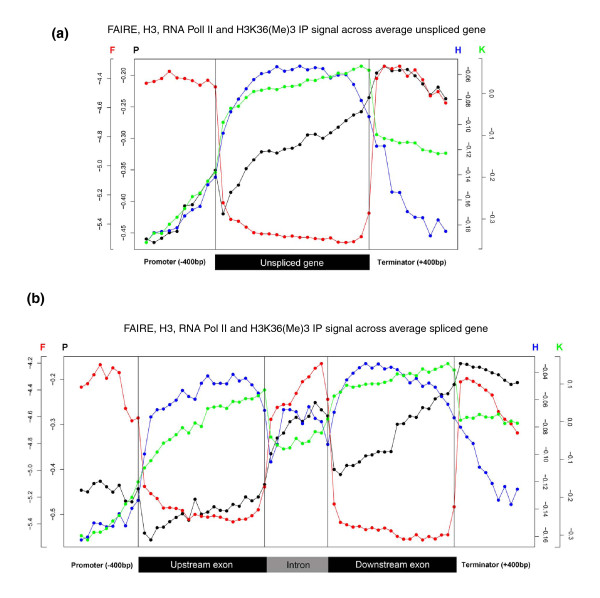
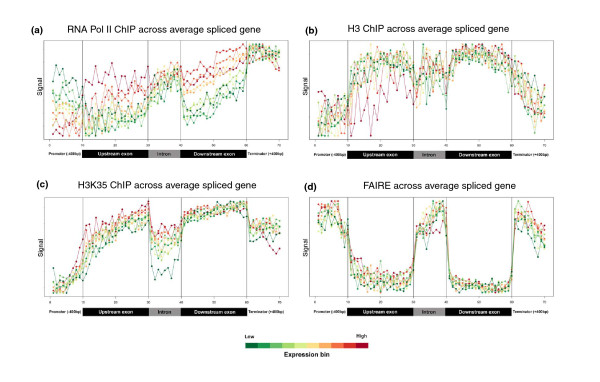
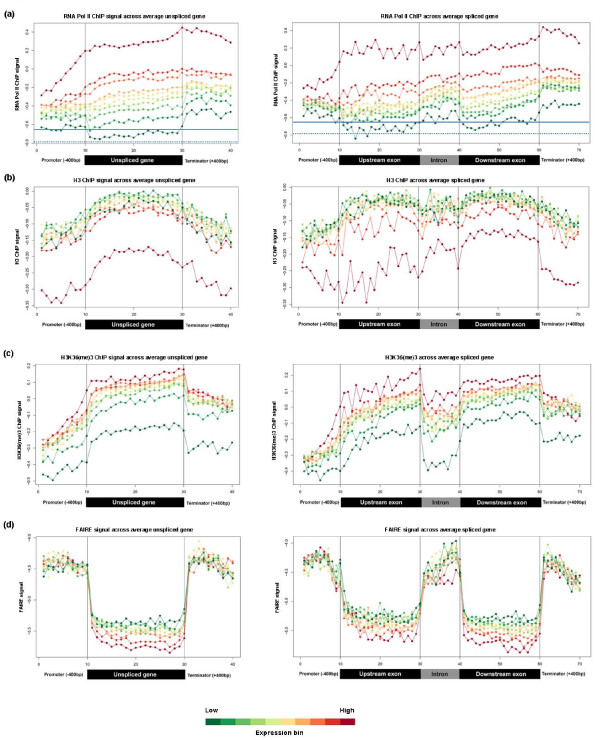
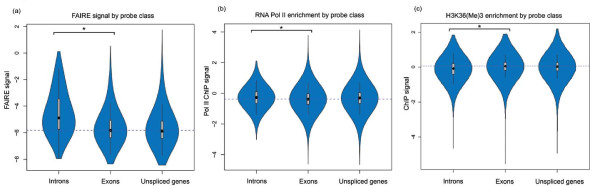
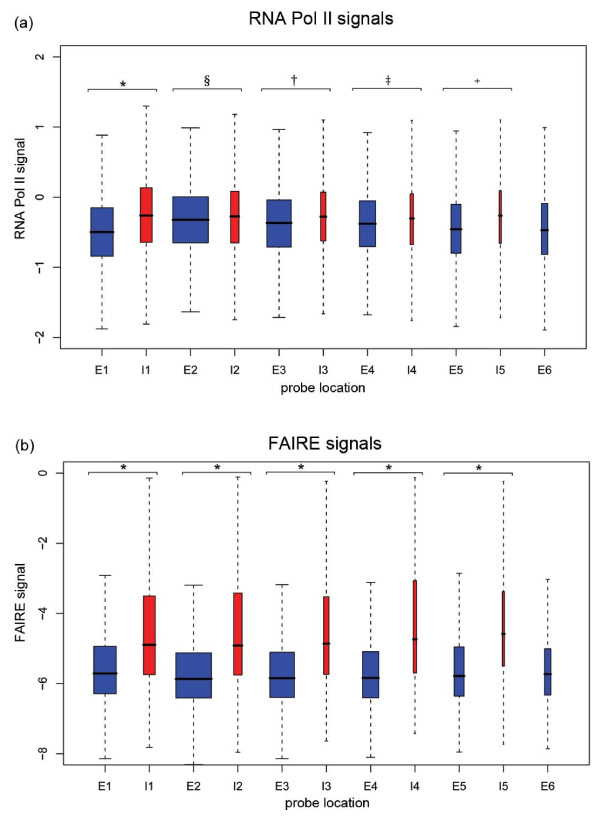
To investigate these functional interdependencies, we determined Pol II occupancy across all genes using high-density tiling arrays. We also performed ChIP-chip on the same array platform to globally map histone H3 and its H3K36me3 modification, complemented by formaldehyde-assisted isolation of regulatory elements (FAIRE). Surprisingly, Pol II occupancy was higher in introns than in exons, and this difference was inversely correlated with gene expression levels at a global level. Moreover, introns showed distinct distributions of histone H3, H3K36me3 and FAIRE signals, similar to those at promoters and terminators. These distinct transcription and chromatin patterns of intronic regions were most pronounced in poorly expressed genes.
Our findings suggest that Pol II accumulates at the 3' ends of introns, leading to substantial transcriptional delays in weakly transcribed genes. We propose that the global relationship between transcription, chromatin remodeling, and splicing may reflect differences in local nuclear environments, with highly expressed genes being associated with abundant processing factors that promote effective intron splicing and transcriptional elongation.
成熟 mRNA 的生成涉及相互关联的过程,包括 RNA 聚合酶 II(Pol II)转录、组蛋白修饰以及通过加帽、内含子剪接和多聚腺苷酸化对前体 mRNA 进行加工。这些过程被认为在空间和时间上是整合的,但目前尚不清楚这些连接如何在染色质模式和转录动力学的全局水平上表现出来。我们试图通过裂殖酵母中的多种全基因组方法来阐明染色质、转录和剪接之间的关系。
为了研究这些功能的相互依存关系,我们使用高密度平铺阵列确定了所有基因中的 Pol II 占有率。我们还在同一阵列平台上进行了组蛋白 H3 及其 H3K36me3 修饰的 ChIP-chip 实验,同时还进行了甲醛辅助分离调控元件(FAIRE)实验。令人惊讶的是,Pol II 占有率在内含子中高于外显子,并且这种差异与全局基因表达水平呈负相关。此外,内含子显示出组蛋白 H3、H3K36me3 和 FAIRE 信号的独特分布,与启动子和终止子相似。在表达水平较低的基因中,内含子区域的这些独特的转录和染色质模式最为明显。
我们的研究结果表明,Pol II 在内含子的 3' 末端积累,导致弱转录基因的转录延迟显著。我们提出,转录、染色质重塑和剪接之间的全局关系可能反映了局部核环境的差异,高表达基因与丰富的加工因子相关,这些因子促进有效的内含子剪接和转录延伸。