Department of Genetics, Harvard Medical School, Boston, Massachusetts 02115, USA.
G3 (Bethesda). 2013 Mar;3(3):573-83. doi: 10.1534/g3.112.005389. Epub 2013 Mar 1.
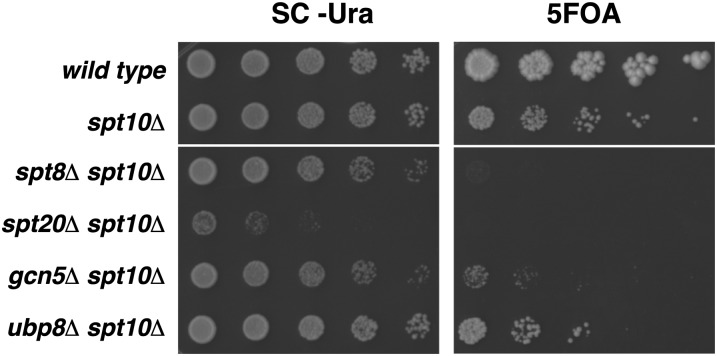
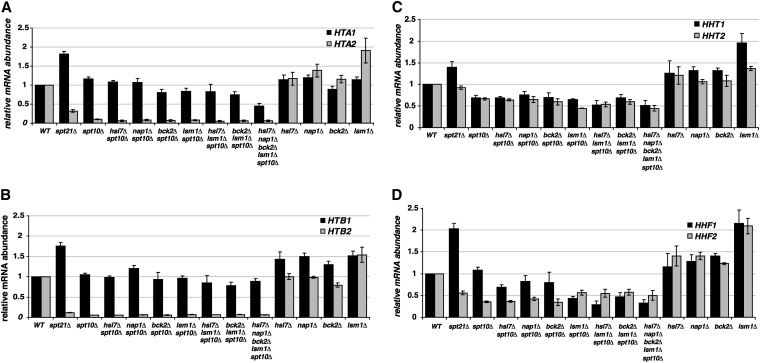
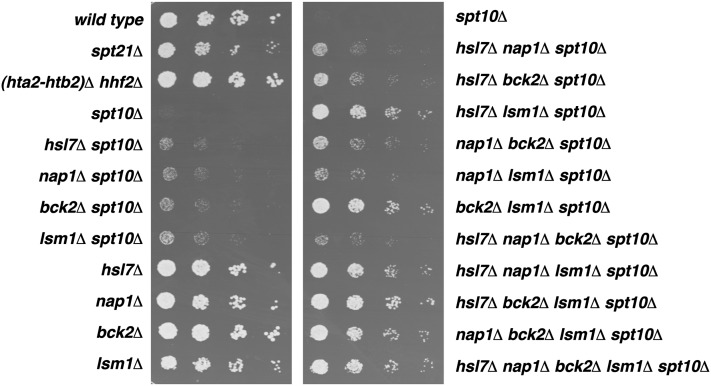
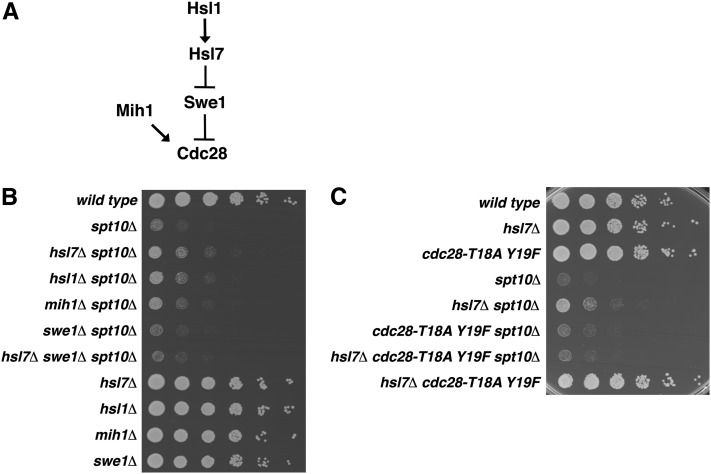
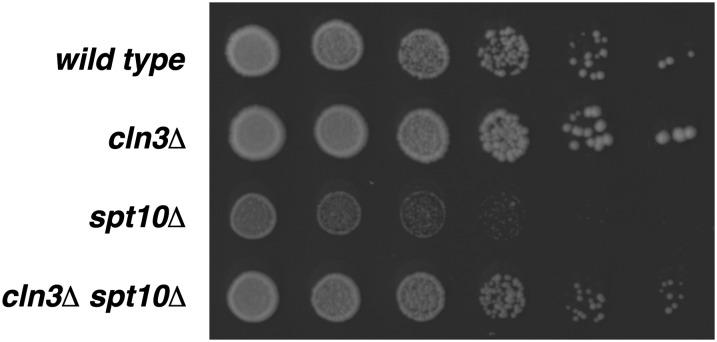
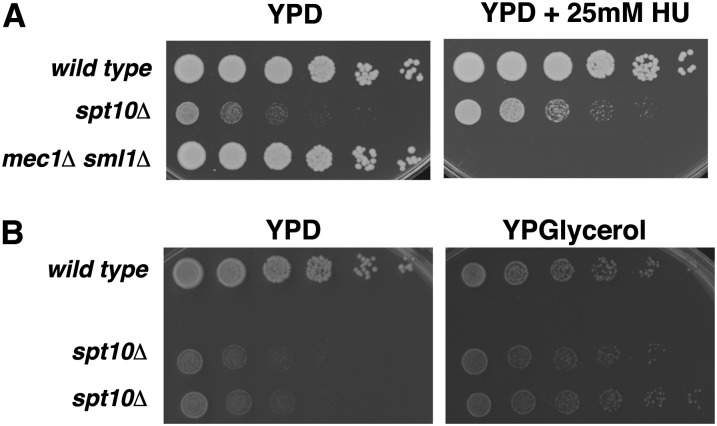
Spt10 is a putative acetyltransferase of Saccharomyces cerevisiae that directly activates the transcription of histone genes. Deletion of SPT10 causes a severe slow growth phenotype, showing that Spt10 is critical for normal cell division. To gain insight into the function of Spt10, we identified mutations that impair or improve the growth of spt10 null (spt10Δ) mutants. Mutations that cause lethality in combination with spt10Δ include particular components of the SAGA complex as well as asf1Δ and hir1Δ. Partial suppressors of the spt10Δ growth defect include mutations that perturb cell-cycle progression through the G1/S transition, S phase, and G2/M. Consistent with these results, slowing of cell-cycle progression by treatment with hydroxyurea or growth on medium containing glycerol as the carbon source also partially suppresses the spt10Δ slow-growth defect. In addition, mutations that impair the Lsm1-7-Pat1 complex, which regulates decapping of polyadenylated mRNAs, also partially suppress the spt10Δ growth defect. Interestingly, suppression of the spt10Δ growth defect is not accompanied by a restoration of normal histone mRNA levels. These findings suggest that Spt10 has multiple roles during cell division.
Spt10 是酿酒酵母中的一种假定乙酰转移酶,它可直接激活组蛋白基因的转录。Spt10 的缺失会导致严重的生长缓慢表型,表明 Spt10 对正常细胞分裂至关重要。为了深入了解 Spt10 的功能,我们鉴定了导致 spt10Δ 缺失突变体生长缺陷或改善其生长的突变。与 spt10Δ 组合导致致死的突变包括 SAGA 复合物的特定成分,以及 asf1Δ 和 hir1Δ。spt10Δ 生长缺陷的部分抑制因子包括通过 G1/S 转换、S 期和 G2/M 期细胞周期进程扰乱的突变。这些结果一致表明,通过羟基脲处理或在含有甘油作为碳源的培养基上生长来减缓细胞周期进程,也可部分抑制 spt10Δ 的生长缓慢缺陷。此外,破坏调节多聚腺苷酸化 mRNA 去帽的 Lsm1-7-Pat1 复合物的突变也部分抑制了 spt10Δ 的生长缺陷。有趣的是,spt10Δ 生长缺陷的抑制并不伴随着正常组蛋白 mRNA 水平的恢复。这些发现表明,Spt10 在细胞分裂过程中有多种作用。