Horitani Masaki, Byer Amanda S, Shisler Krista A, Chandra Tilak, Broderick Joan B, Hoffman Brian M
†Department of Chemistry, Northwestern University, Evanston, Illinois 60208, United States.
‡Department of Chemistry and Biochemistry, Montana State University, Bozeman, Montana 59717, United States.
J Am Chem Soc. 2015 Jun 10;137(22):7111-21. doi: 10.1021/jacs.5b00498. Epub 2015 May 19.
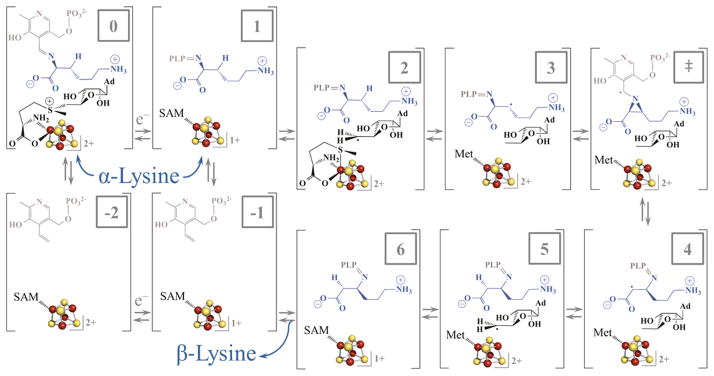
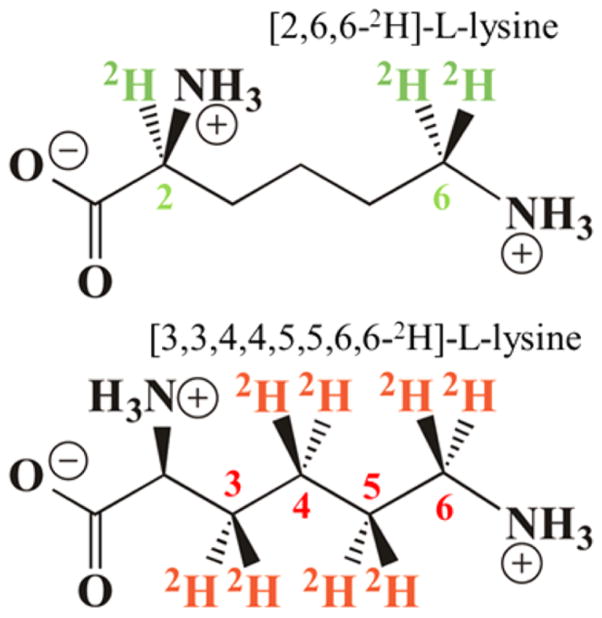
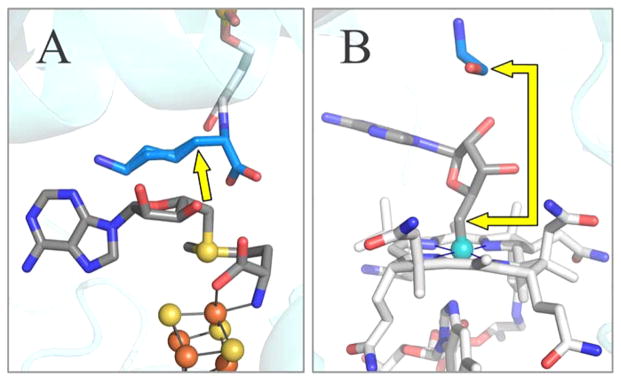
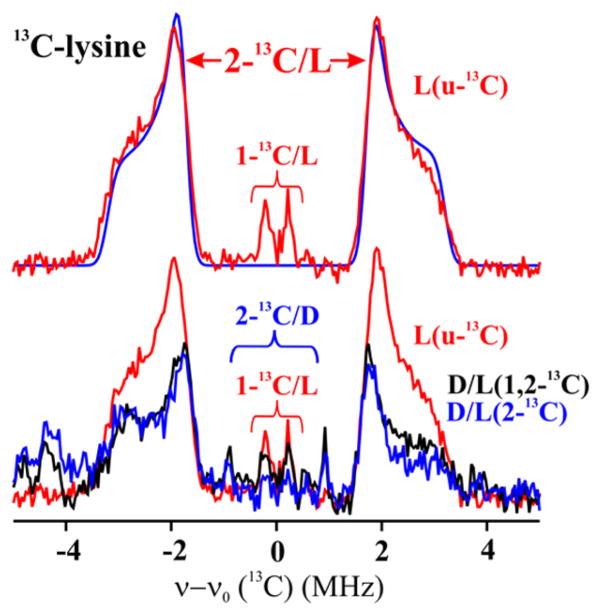
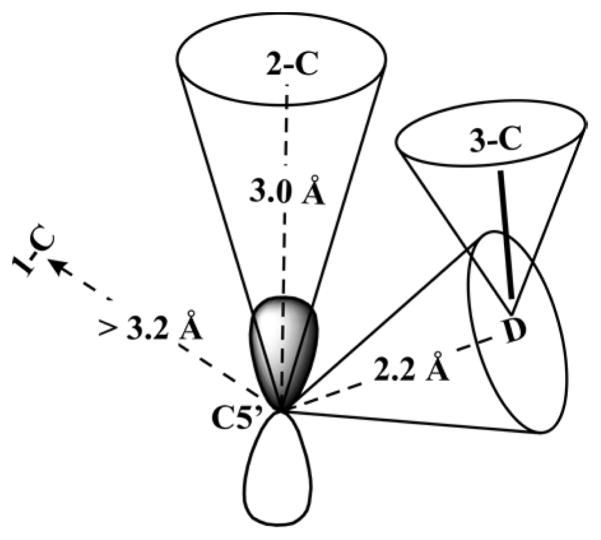
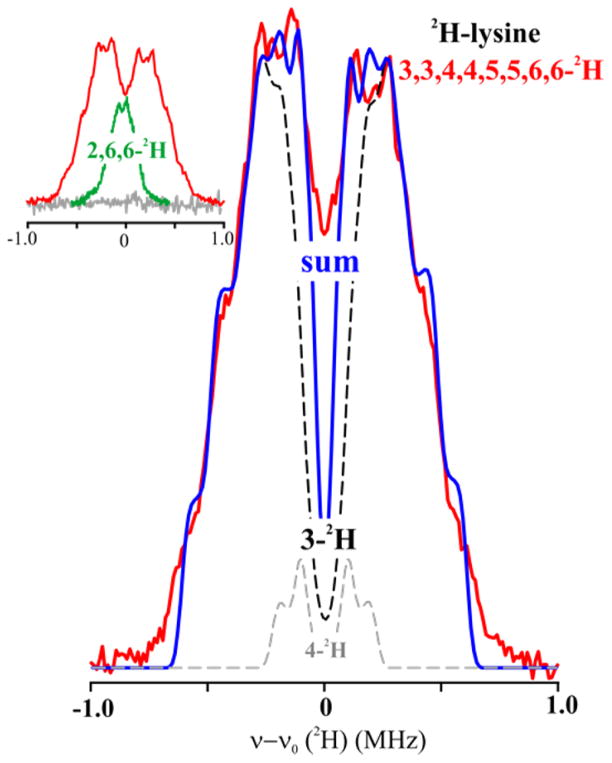
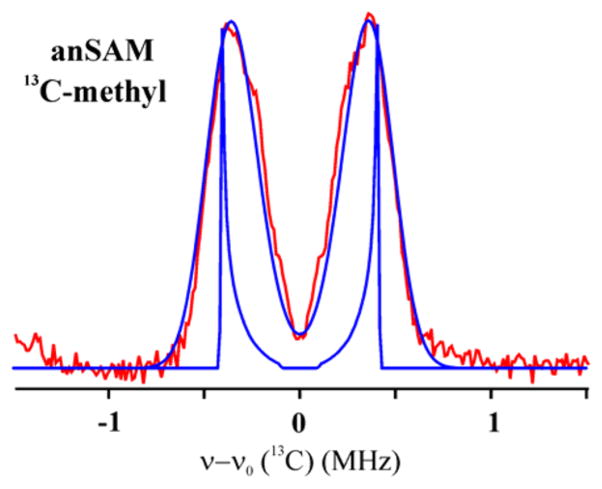
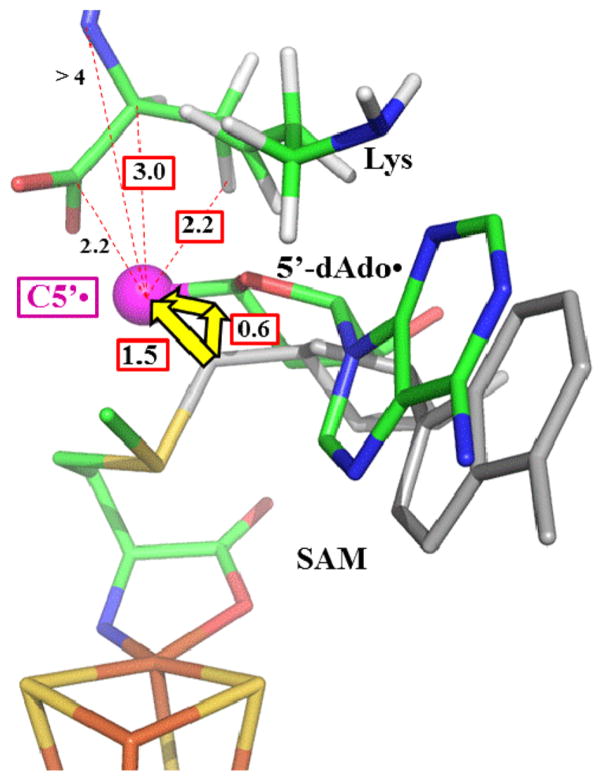
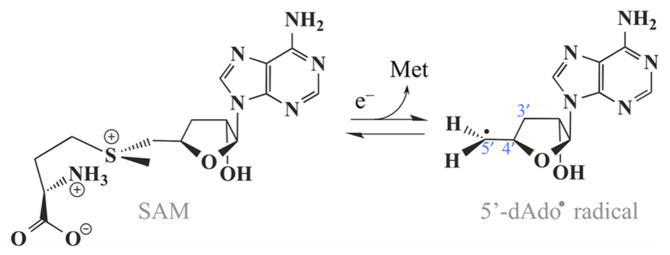
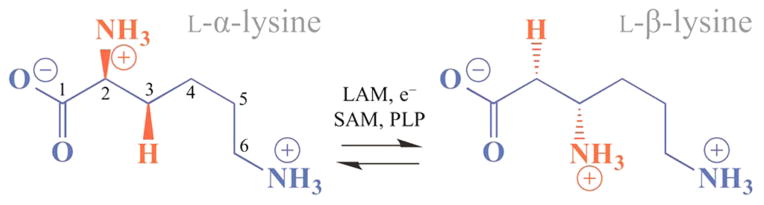
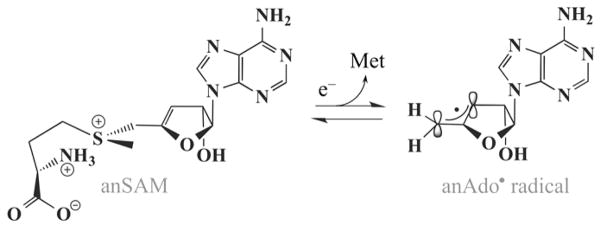
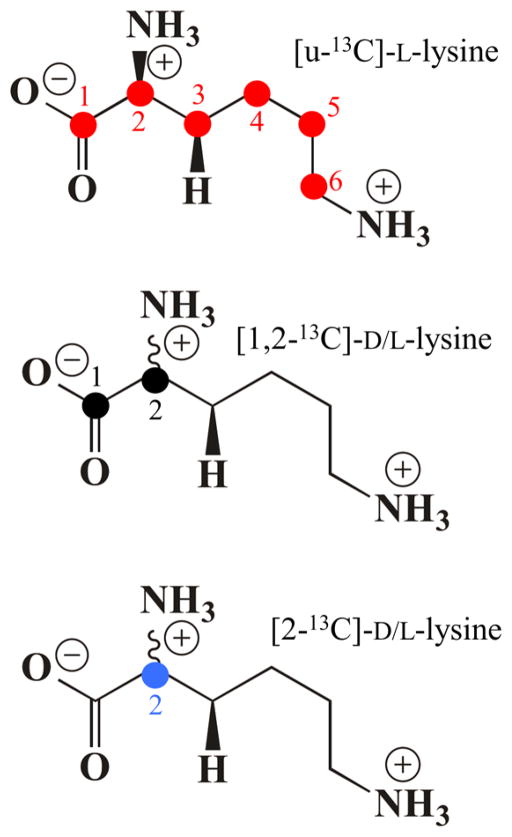
Lysine 2,3-aminomutase (LAM) is a radical S-adenosyl-L-methionine (SAM) enzyme and, like other members of this superfamily, LAM utilizes radical-generating machinery comprising SAM anchored to the unique Fe of a [4Fe-4S] cluster via a classical five-membered N,O chelate ring. Catalysis is initiated by reductive cleavage of the SAM S-C5' bond, which creates the highly reactive 5'-deoxyadenosyl radical (5'-dAdo•), the same radical generated by homolytic Co-C bond cleavage in B12 radical enzymes. The SAM surrogate S-3',4'-anhydroadenosyl-L-methionine (anSAM) can replace SAM as a cofactor in the isomerization of L-α-lysine to L-β-lysine by LAM, via the stable allylic anhydroadenosyl radical (anAdo•). Here electron nuclear double resonance (ENDOR) spectroscopy of the anAdo• radical in the presence of (13)C, (2)H, and (15)N-labeled lysine completes the picture of how the active site of LAM from Clostridium subterminale SB4 "tames" the 5'-dAdo• radical, preventing it from carrying out harmful side reactions: this "free radical" in LAM is never free. The low steric demands of the radical-generating [4Fe-4S]/SAM construct allow the substrate target to bind adjacent to the S-C5' bond, thereby enabling the 5'-dAdo• radical created by cleavage of this bond to react with its partners by undergoing small motions, ∼0.6 Å toward the target and ∼1.5 Å overall, that are controlled by tight van der Waals contact with its partners. We suggest that the accessibility to substrate and ready control of the reactive C5' radical, with "van der Waals control" of small motions throughout the catalytic cycle, is common within the radical SAM enzyme superfamily and is a major reason why these enzymes are the preferred means of initiating radical reactions in nature.
赖氨酸2,3-氨基变位酶(LAM)是一种自由基S-腺苷-L-甲硫氨酸(SAM)酶,与该超家族的其他成员一样,LAM利用自由基生成机制,该机制包括通过经典的五元N,O螯合环锚定在[4Fe-4S]簇独特铁原子上的SAM。催化作用由SAM的S-C5'键的还原裂解引发,这产生了高反应性的5'-脱氧腺苷自由基(5'-dAdo•),与B12自由基酶中通过均裂Co-C键裂解产生的自由基相同。SAM替代物S-3',4'-脱水腺苷-L-甲硫氨酸(anSAM)可以替代SAM作为辅因子,通过稳定的烯丙基脱水腺苷自由基(anAdo•),在LAM催化L-α-赖氨酸异构化为L-β-赖氨酸的过程中发挥作用。在这里,在存在(13)C、(2)H和(15)N标记的赖氨酸的情况下,对anAdo•自由基进行电子核双共振(ENDOR)光谱分析,完善了来自Clostridium subterminale SB4的LAM活性位点如何“驯服”5'-dAdo•自由基、防止其进行有害副反应的图景:LAM中的这个“自由基”从未自由过。产生自由基的[4Fe-4S]/SAM结构的低空间需求允许底物靶点结合在S-C5'键附近,从而使通过该键裂解产生的5'-dAdo•自由基能够通过向靶点进行约0.6 Å、总体约1.5 Å的小移动与其伙伴发生反应,这些移动由与伙伴的紧密范德华接触控制。我们认为,底物的可及性以及对反应性C5'自由基的随时控制,以及在整个催化循环中对小移动的“范德华控制”,在自由基SAM酶超家族中是常见的,并且是这些酶成为自然界中引发自由基反应的首选方式的主要原因。