Scarabelli Guido, Soppina Virupakshi, Yao Xin-Qiu, Atherton Joseph, Moores Carolyn A, Verhey Kristen J, Grant Barry J
Department of Computational Medicine and Bioinformatics, University of Michigan Medical School, Ann Arbor, Michigan.
Department of Cell and Developmental Biology, University of Michigan Medical School, Ann Arbor, Michigan.
Biophys J. 2015 Oct 20;109(8):1537-40. doi: 10.1016/j.bpj.2015.08.027.
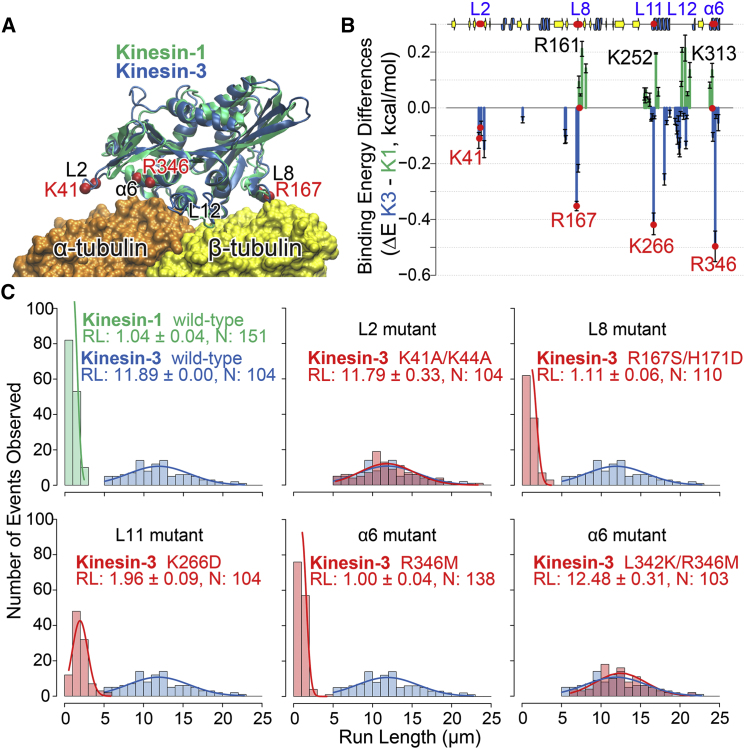
Kinesin superfamily members play important roles in many diverse cellular processes, including cell motility, cell division, intracellular transport, and regulation of the microtubule cytoskeleton. How the properties of the family-defining motor domain of distinct kinesins are tailored to their different cellular roles remains largely unknown. Here, we employed molecular-dynamics simulations coupled with energetic calculations to infer the family-specific interactions of kinesin-1 and kinesin-3 motor domains with microtubules in different nucleotide states. We then used experimental mutagenesis and single-molecule motility assays to further assess the predicted residue-wise determinants of distinct kinesin-microtubule binding properties. Collectively, our results identify residues in the L8, L11, and α6 regions that contribute to family-specific microtubule interactions and whose mutation affects motor-microtubule complex stability and processive motility (the ability of an individual motor to take multiple steps along its microtubule filament). In particular, substitutions of prominent kinesin-3 residues with those found in kinesin-1, namely, R167S/H171D, K266D, and R346M, were found to decrease kinesin-3 processivity 10-fold and thus approach kinesin-1 levels.
驱动蛋白超家族成员在许多不同的细胞过程中发挥着重要作用,包括细胞运动、细胞分裂、细胞内运输以及微管细胞骨架的调节。不同驱动蛋白的家族定义运动结构域的特性是如何根据其不同的细胞作用进行调整的,目前仍 largely 未知。在这里,我们采用分子动力学模拟结合能量计算,来推断驱动蛋白 -1 和驱动蛋白 -3 运动结构域在不同核苷酸状态下与微管的家族特异性相互作用。然后,我们使用实验性诱变和单分子运动分析,进一步评估预测的不同驱动蛋白 - 微管结合特性的逐个残基决定因素。总的来说,我们的结果确定了 L8、L11 和 α6 区域中的残基,这些残基有助于家族特异性微管相互作用,并且其突变会影响运动蛋白 - 微管复合物的稳定性和持续运动性(单个运动蛋白沿着其微管丝采取多个步骤的能力)。特别是,发现用驱动蛋白 -1 中发现的残基替代驱动蛋白 -3 的突出残基,即 R167S/H171D、K266D 和 R346M,会使驱动蛋白 -3 的持续运动性降低 10 倍,从而接近驱动蛋白 -1 的水平。