Lu Wen-Juan, Tseng Scheffer C G, Chen Shuangling, Tighe Sean, Zhang Yuan, Liu Xin, Chen Szu-Yu, Su Chen-Wei, Zhu Ying-Ting
Department of Ophthalmology, Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China.
R&D Department, Tissue Tech, Inc., Ocular Surface Research &Education Foundation, Miami, FL, USA.
Sci Rep. 2016 Oct 14;6:35166. doi: 10.1038/srep35166.
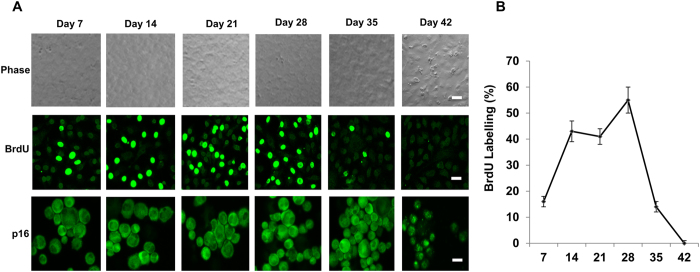
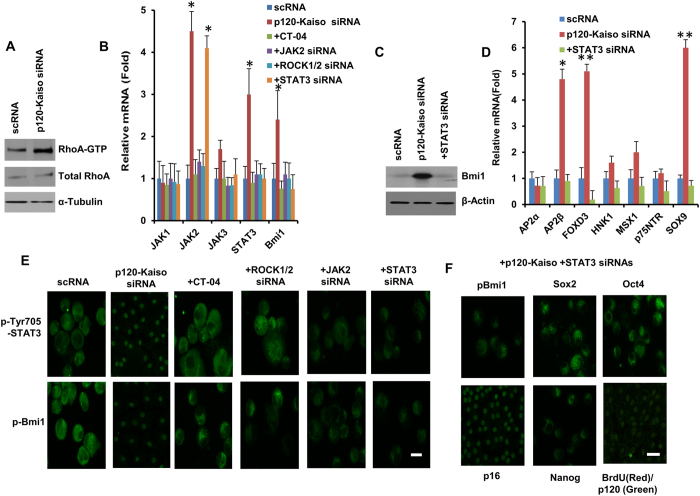
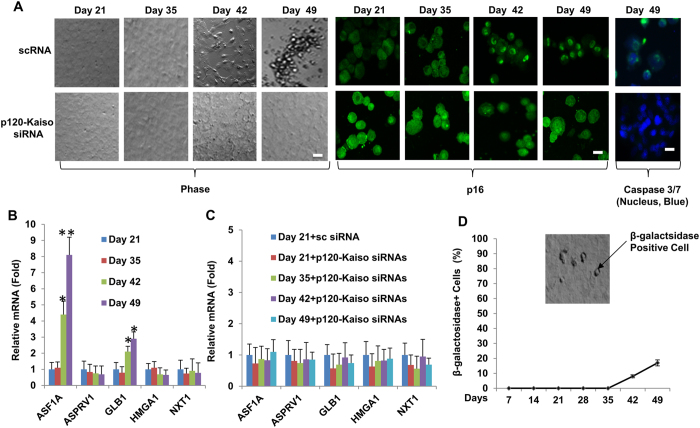
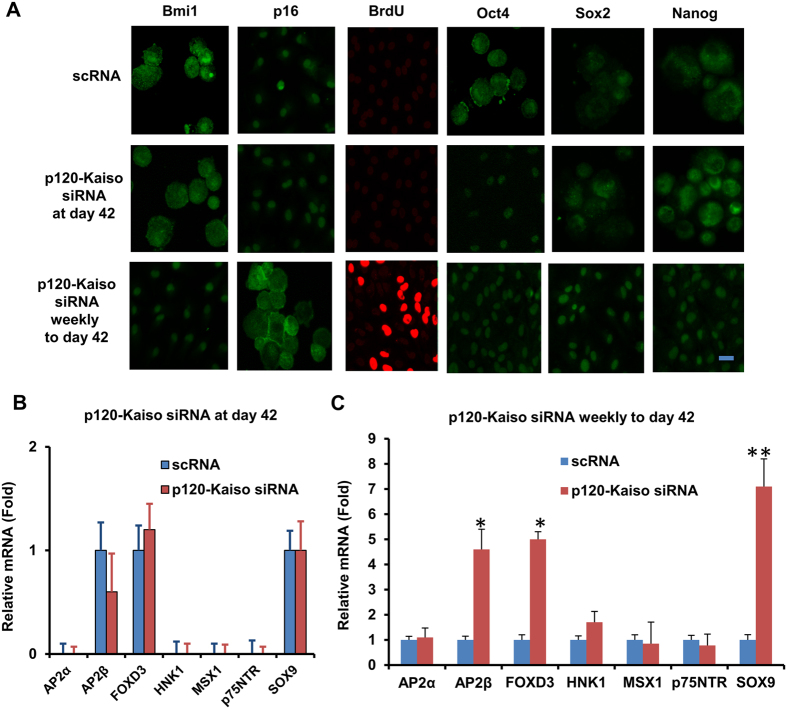
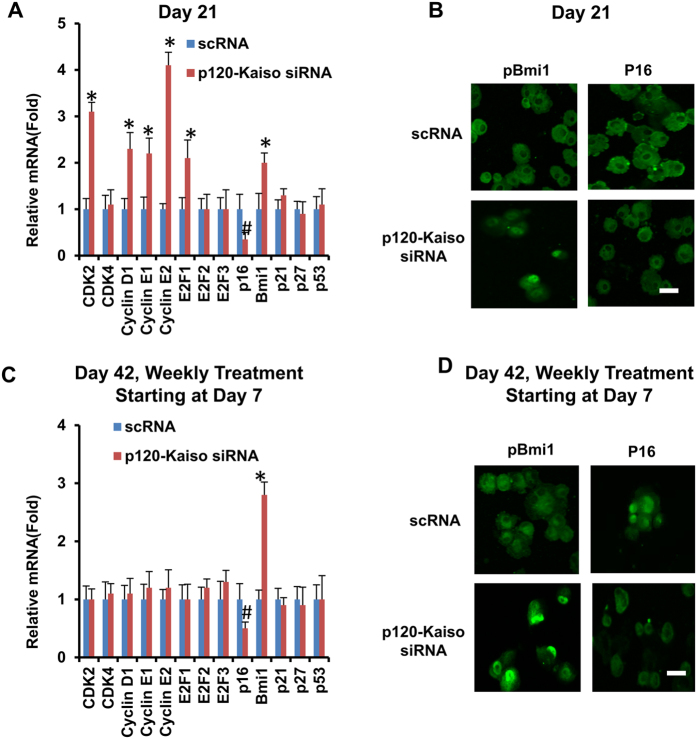
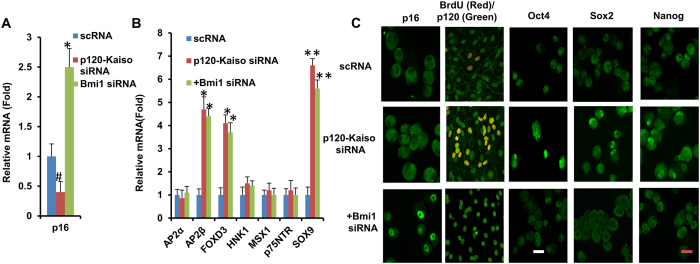
Human corneal endothelial cells (HCECs) have limited proliferative capacity due to "contact-inhibition" at G1 phase. Such contact-inhibition can be delayed from Day 21 to Day 42 by switching EGF-containing SHEM to LIF/bFGF-containing MESCM through transient activation of LIF-JAK1-STAT3 signaling that delays eventual nuclear translocation of p16. Using the latter system, we have reported a novel tissue engineering technique by implementing 5 weekly knockdowns with p120 catenin (p120) and Kaiso siRNAs since Day 7 to achieve effective expansion of HCEC monolayers to a transplantable size with a normal HCEC density, through reprogramming of HCECs into neural crest progenitors by activating p120-Kaiso-RhoA-ROCK-canonical BMP signaling. Herein, we noted that a single knockdown with p120-Kaiso siRNAs at Day 42 failed to achieve such reprogramming when contact inhibition transitioned to senescence with nuclear translocation of p16. In contrast, 5 weekly knockdowns with p120-Kaiso siRNAs since Day 7 precluded senescence mediated by p16 by inducing nuclear translocation of Bmi1 because of sustained activation of JAK2-STAT3 signaling downstream of p120-Kaiso-RhoA-ROCK signaling. STAT3 or Bmi1 siRNA impeded nuclear exclusion of p16 and suppressed the reprogramming induced by p120-Kaiso siRNAs, suggesting that another important engineering strategy of HCEC lies in prevention of senescence mediated by nuclear translocation of p16.
人角膜内皮细胞(HCECs)由于在G1期的“接触抑制”而增殖能力有限。通过短暂激活LIF-JAK1-STAT3信号通路,将含表皮生长因子(EGF)的SHEM换成含白血病抑制因子(LIF)/碱性成纤维细胞生长因子(bFGF)的MESCM,可将这种接触抑制从第21天推迟到第42天,该信号通路可延迟p16最终的核转位。利用后一种体系,我们报道了一种新的组织工程技术,即从第7天开始每周用p120连环蛋白(p120)和Kaiso小干扰RNA(siRNAs)进行5次敲低,通过激活p120-Kaiso-RhoA-ROCK-经典骨形态发生蛋白(BMP)信号通路将HCECs重编程为神经嵴祖细胞,从而有效将HCEC单层扩展到可移植大小并维持正常的HCEC密度。在此,我们注意到,当接触抑制随着p16的核转位转变为衰老时,在第42天用p120-Kaiso siRNAs进行单次敲低无法实现这种重编程。相反,从第7天开始每周用p120-Kaiso siRNAs进行5次敲低,由于p120-Kaiso-RhoA-ROCK信号下游的JAK2-STAT3信号通路持续激活,诱导Bmi1核转位,从而阻止了由p16介导的衰老。STAT3或Bmi1 siRNA阻碍了p16的核排除,并抑制了p120-Kaiso siRNAs诱导的重编程,这表明HCEC的另一个重要工程策略在于预防由p16核转位介导的衰老。