Nokin Marie-Julie, Durieux Florence, Peixoto Paul, Chiavarina Barbara, Peulen Olivier, Blomme Arnaud, Turtoi Andrei, Costanza Brunella, Smargiasso Nicolas, Baiwir Dominique, Scheijen Jean L, Schalkwijk Casper G, Leenders Justine, De Tullio Pascal, Bianchi Elettra, Thiry Marc, Uchida Koji, Spiegel David A, Cochrane James R, Hutton Craig A, De Pauw Edwin, Delvenne Philippe, Belpomme Dominique, Castronovo Vincent, Bellahcène Akeila
Metastasis Research Laboratory, GIGA-CANCER, University of Liège, Liège, Belgium.
Mass Spectrometry Laboratory, GIGA-Systems Biology and Chemical Biology, University of Liège, Liège, Belgium.
Elife. 2016 Oct 19;5:e19375. doi: 10.7554/eLife.19375.
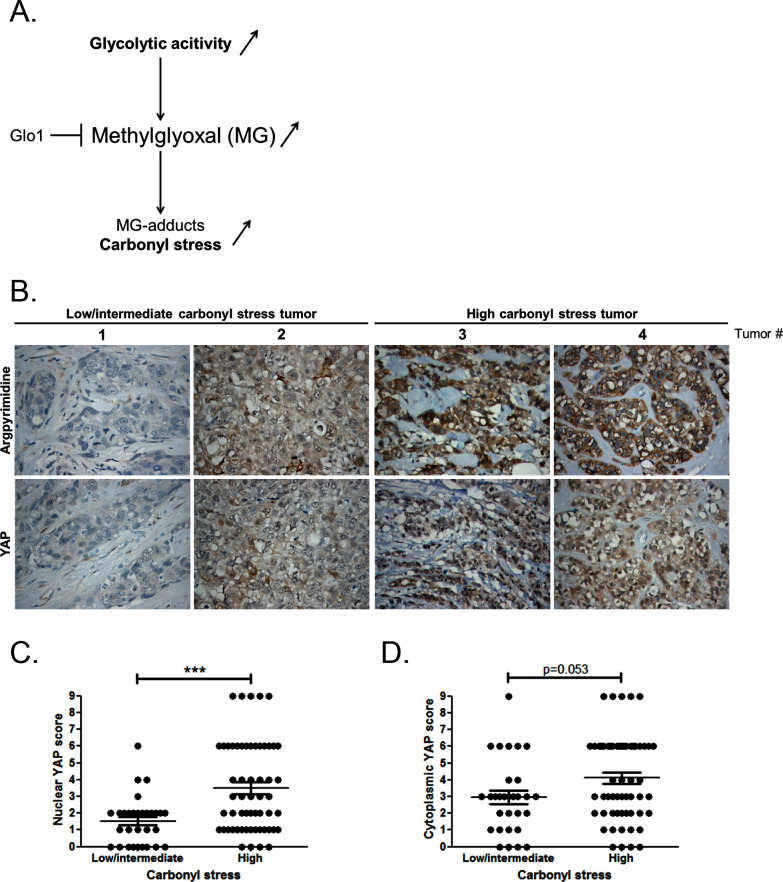
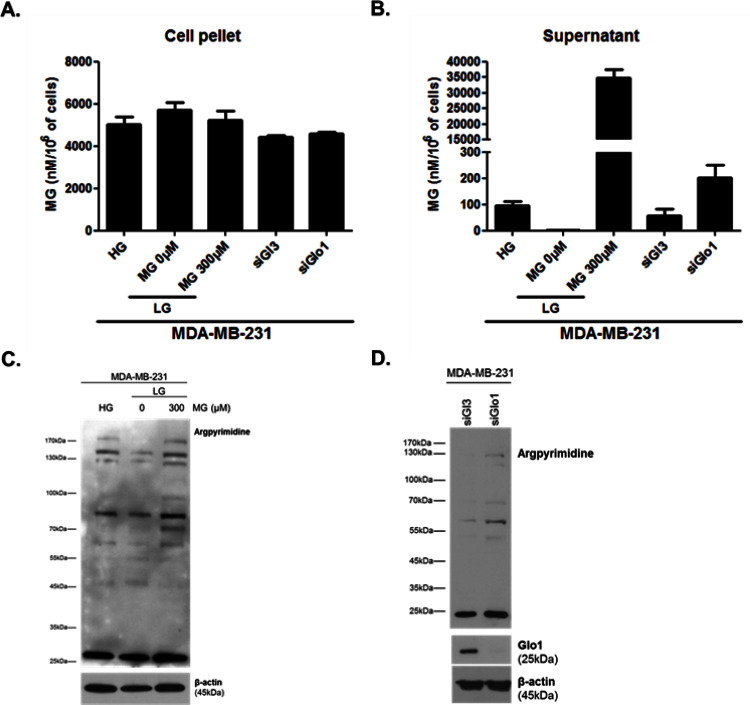
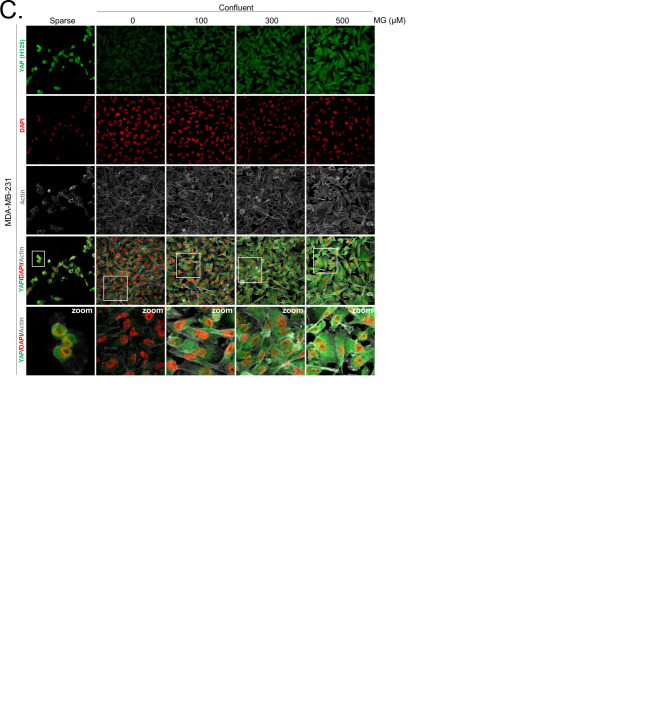
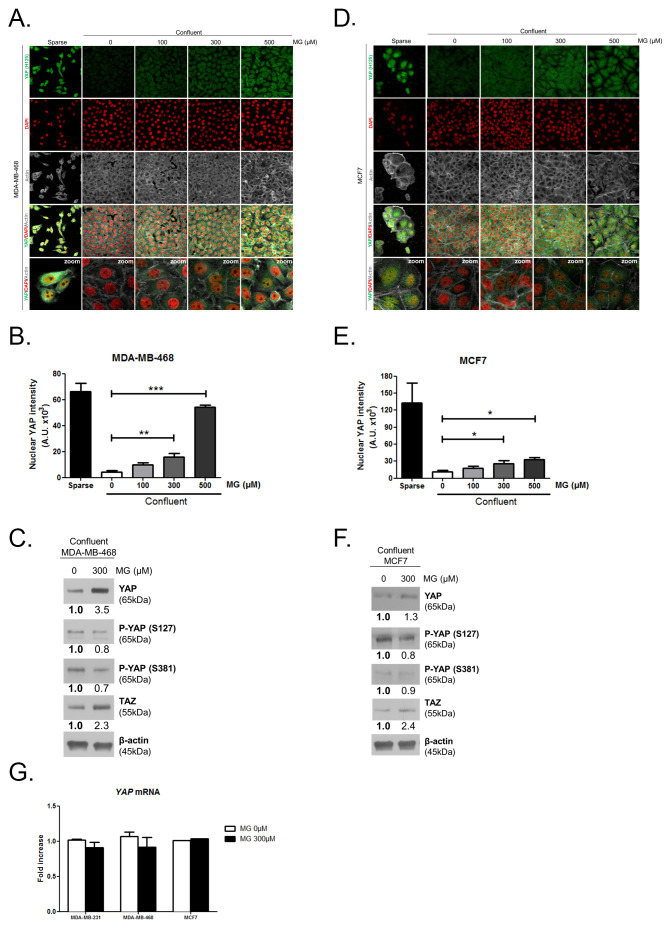
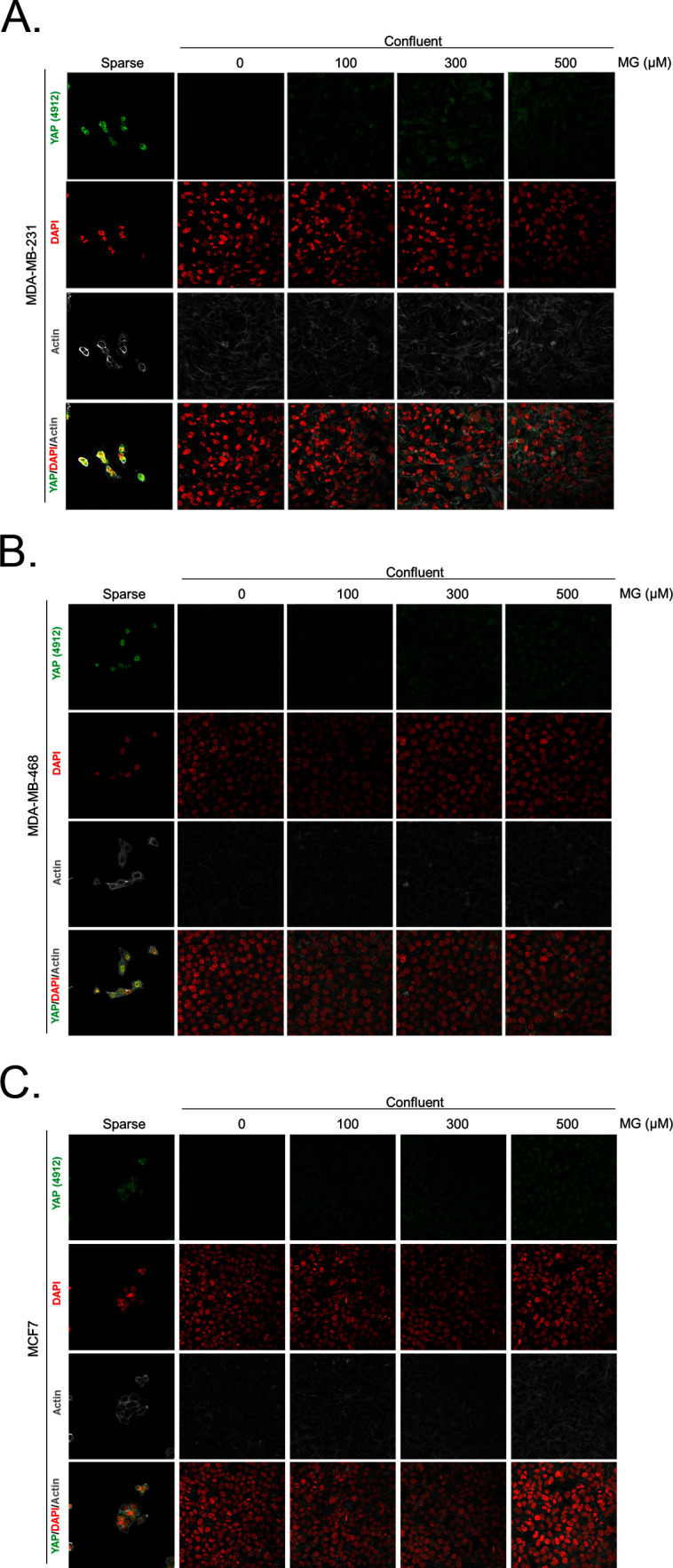
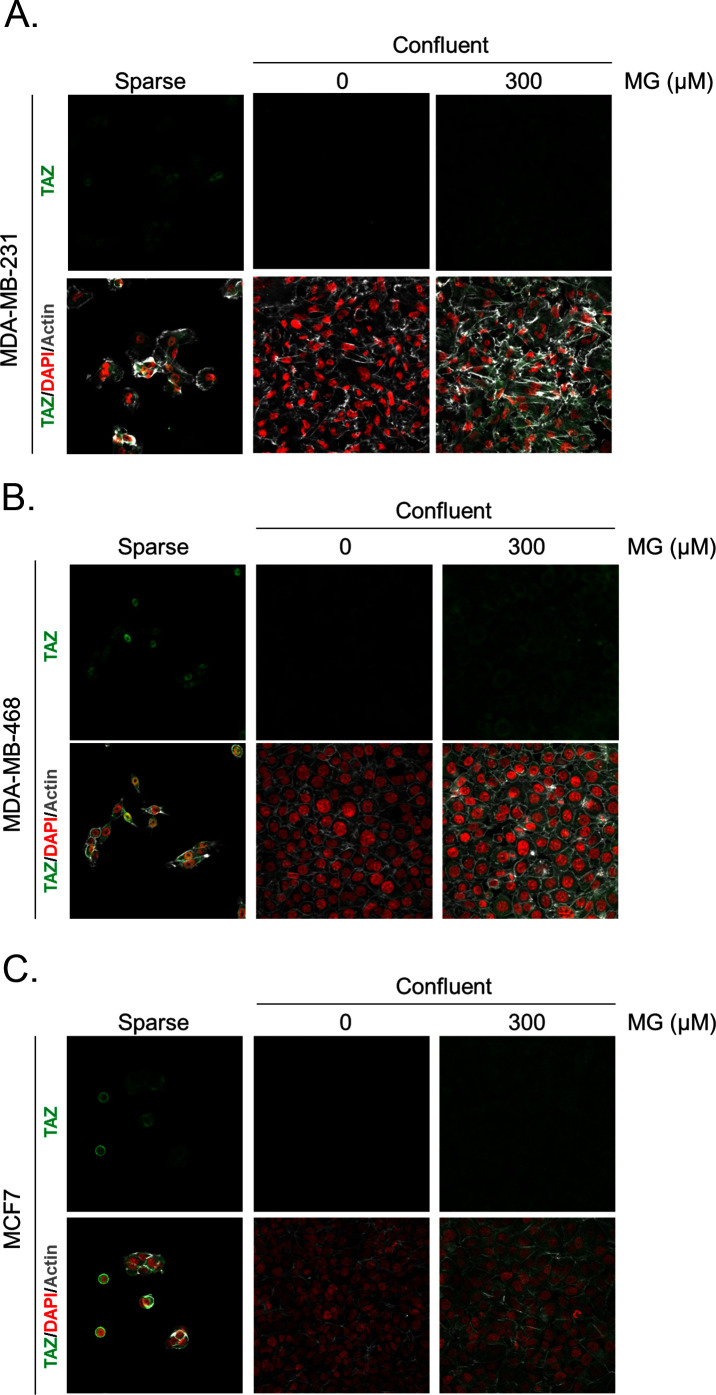
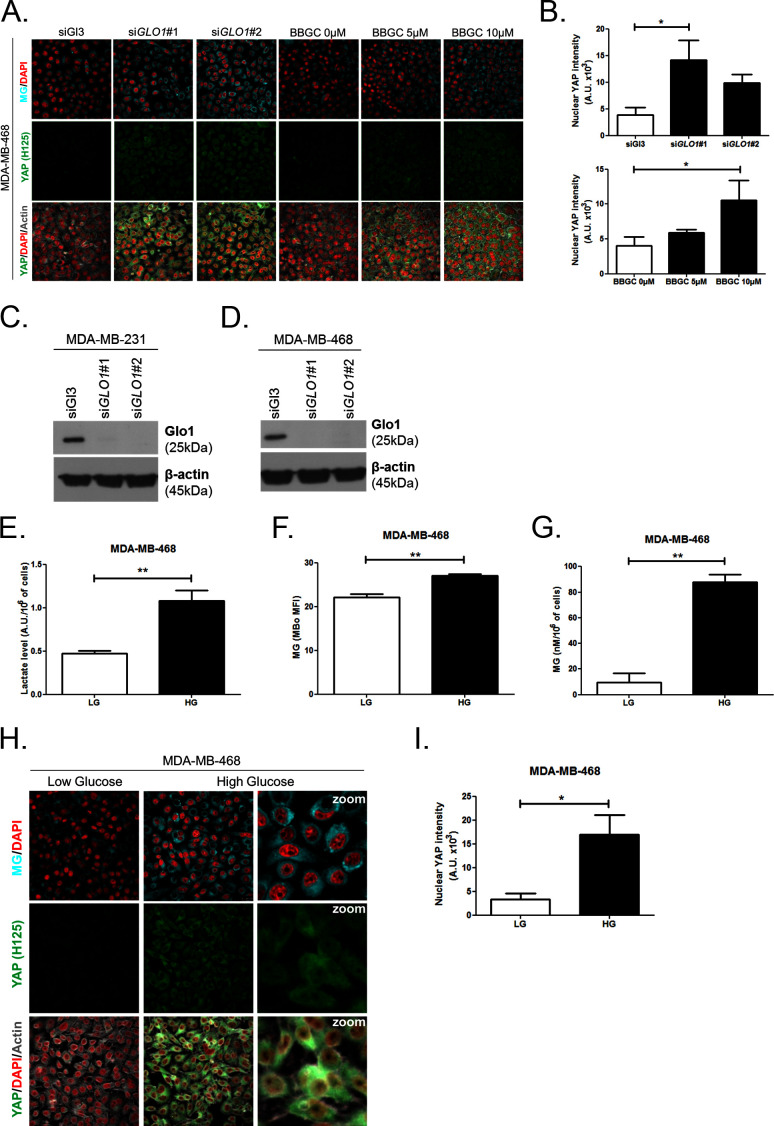
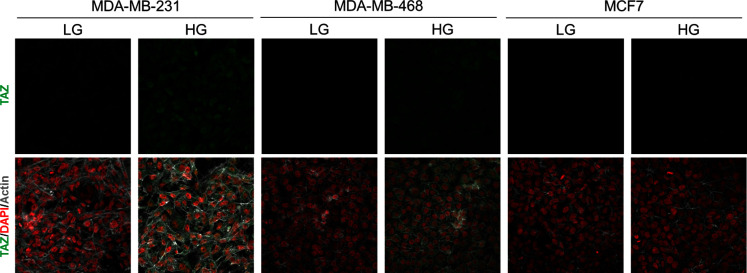
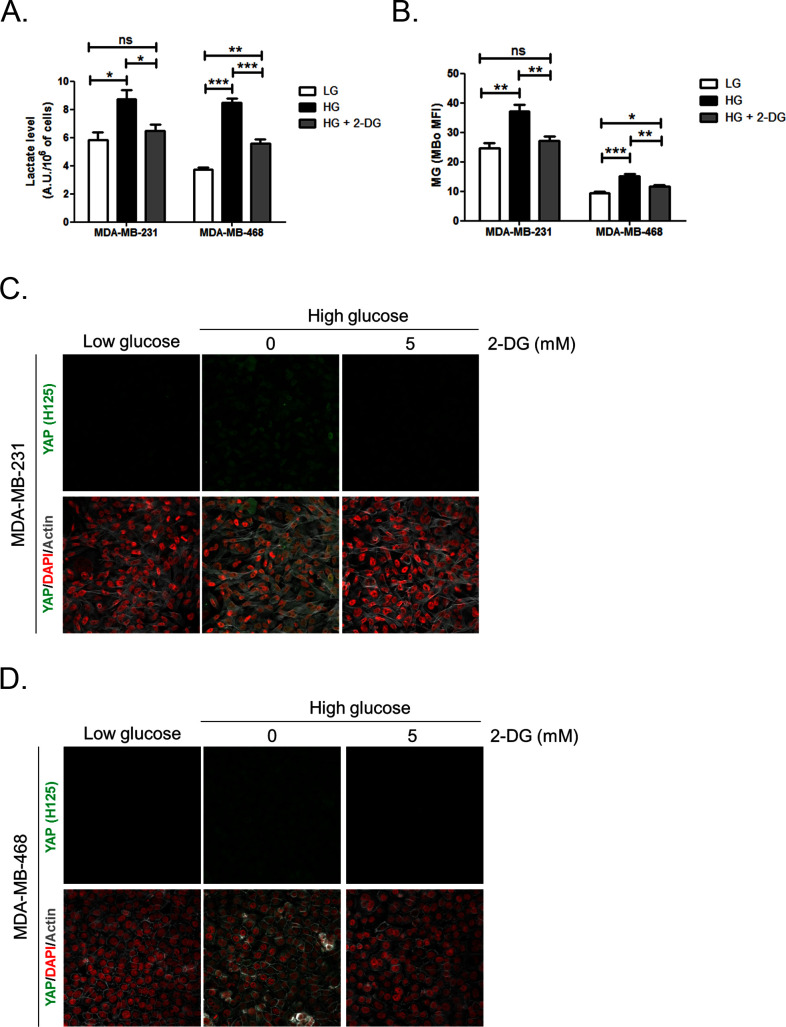
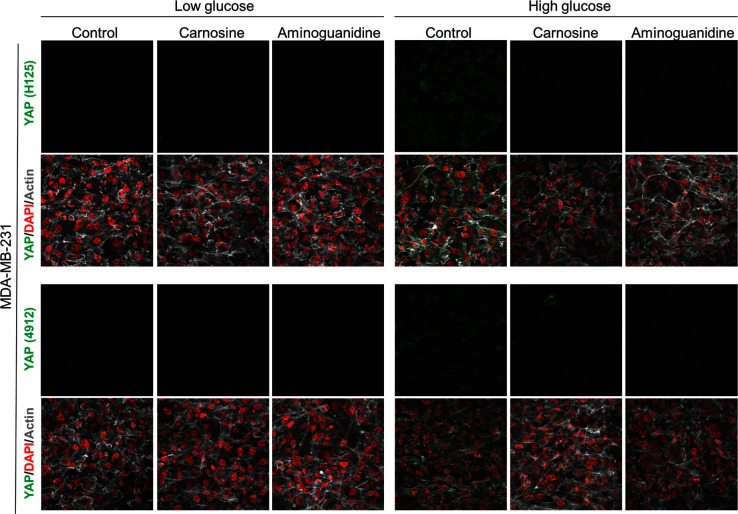
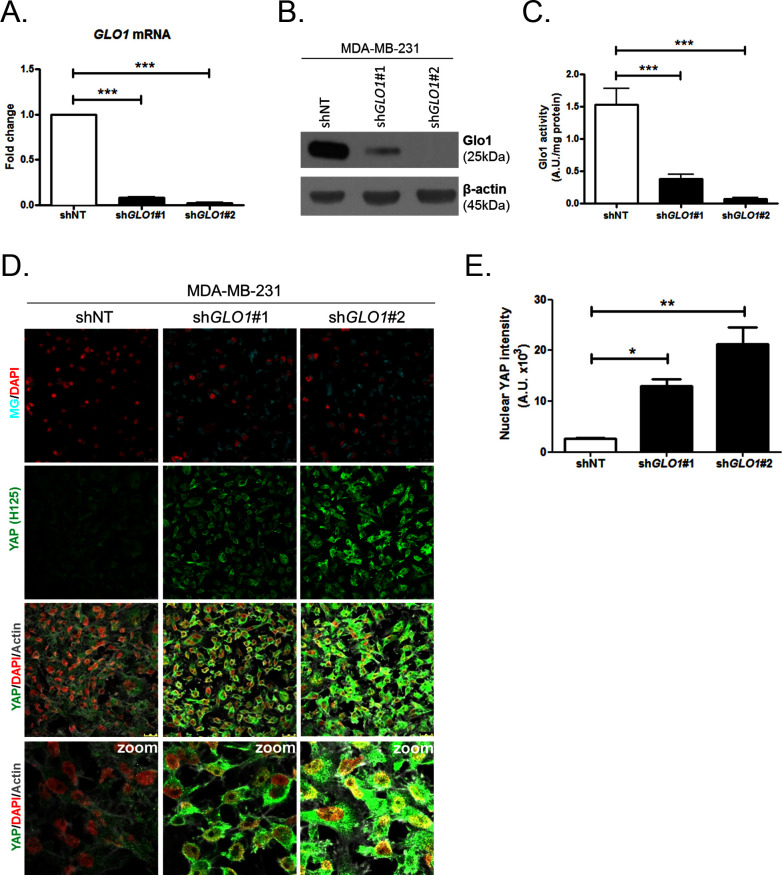
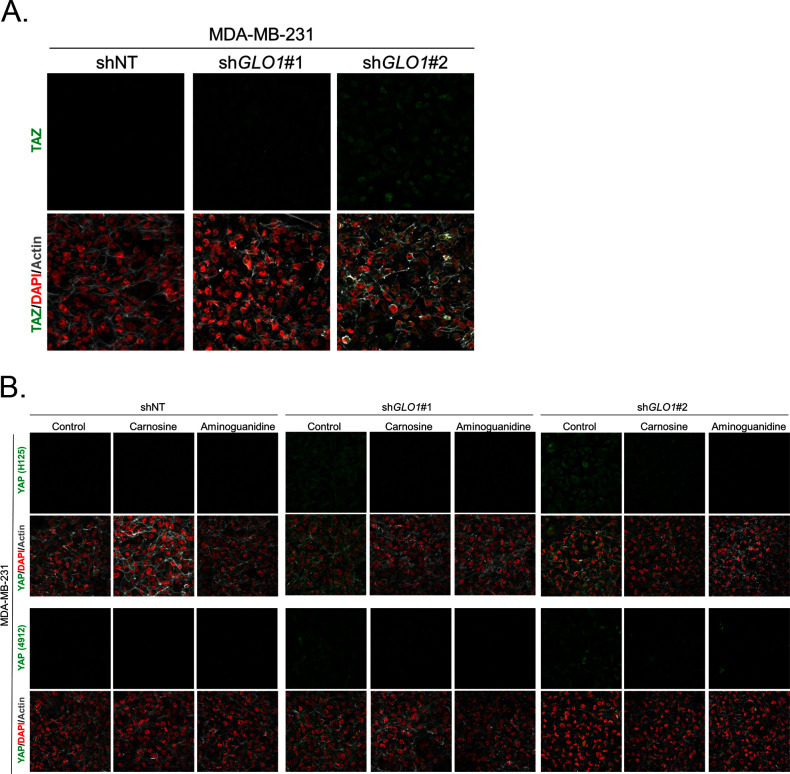
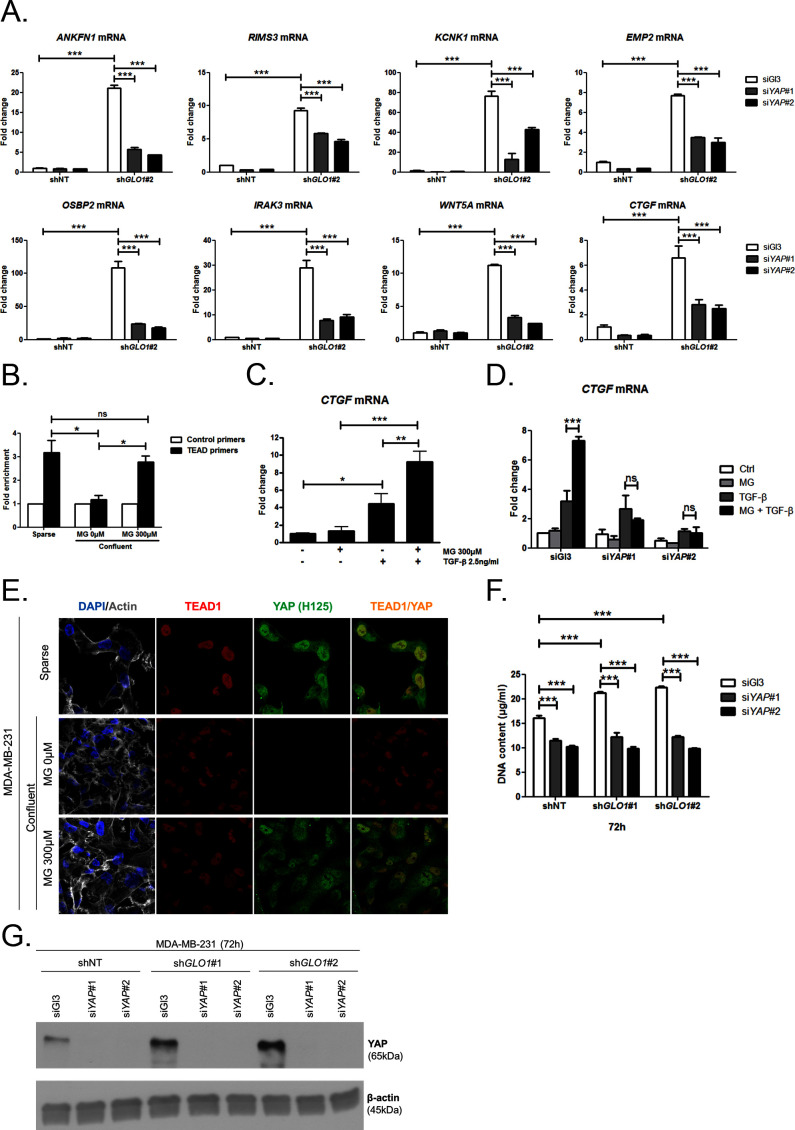
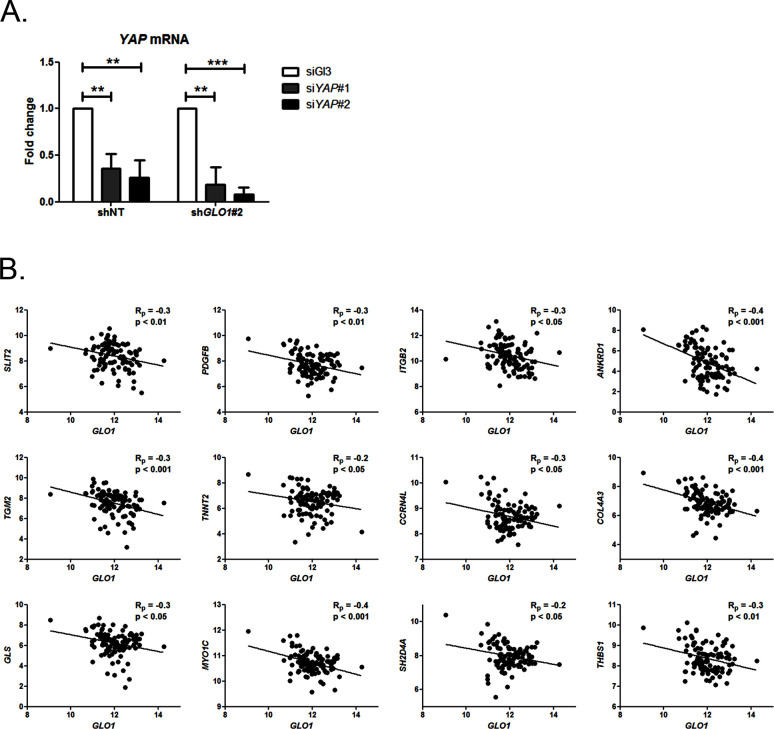
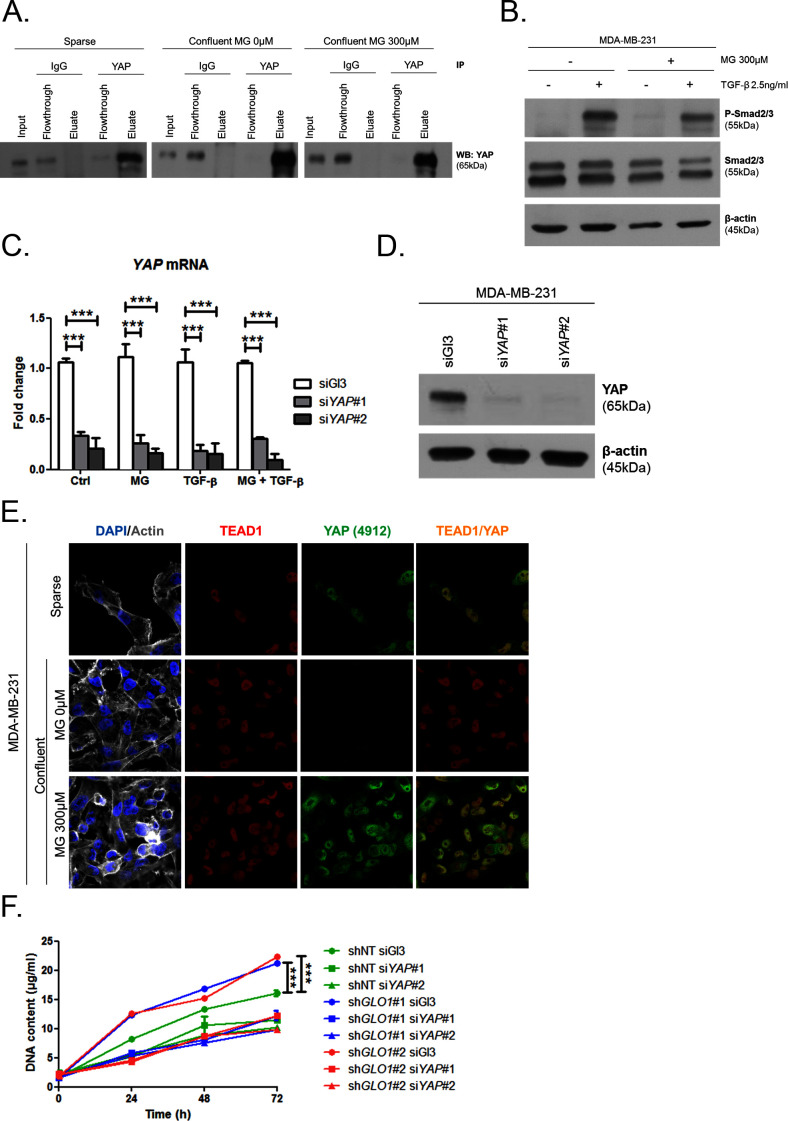
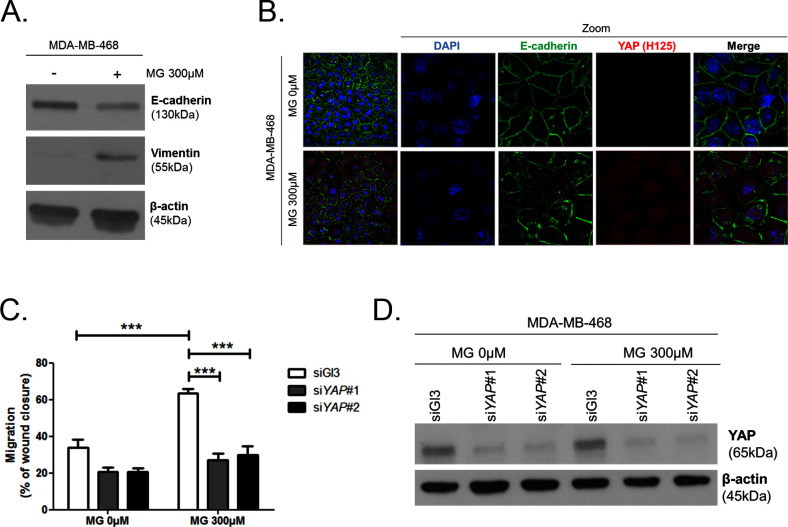
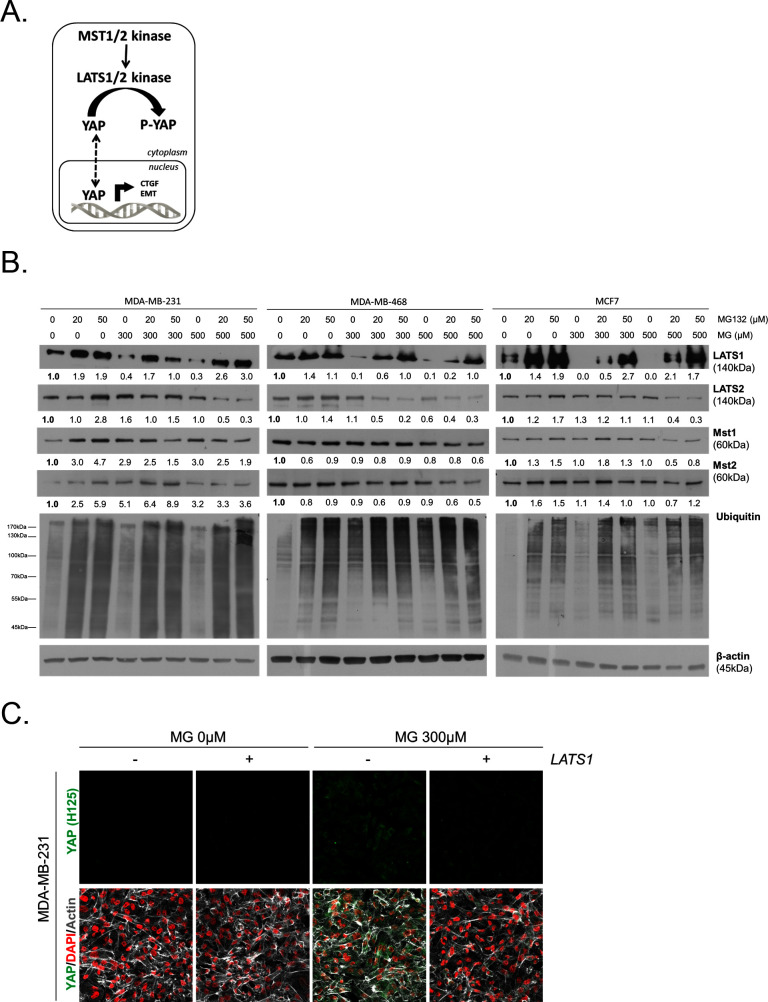
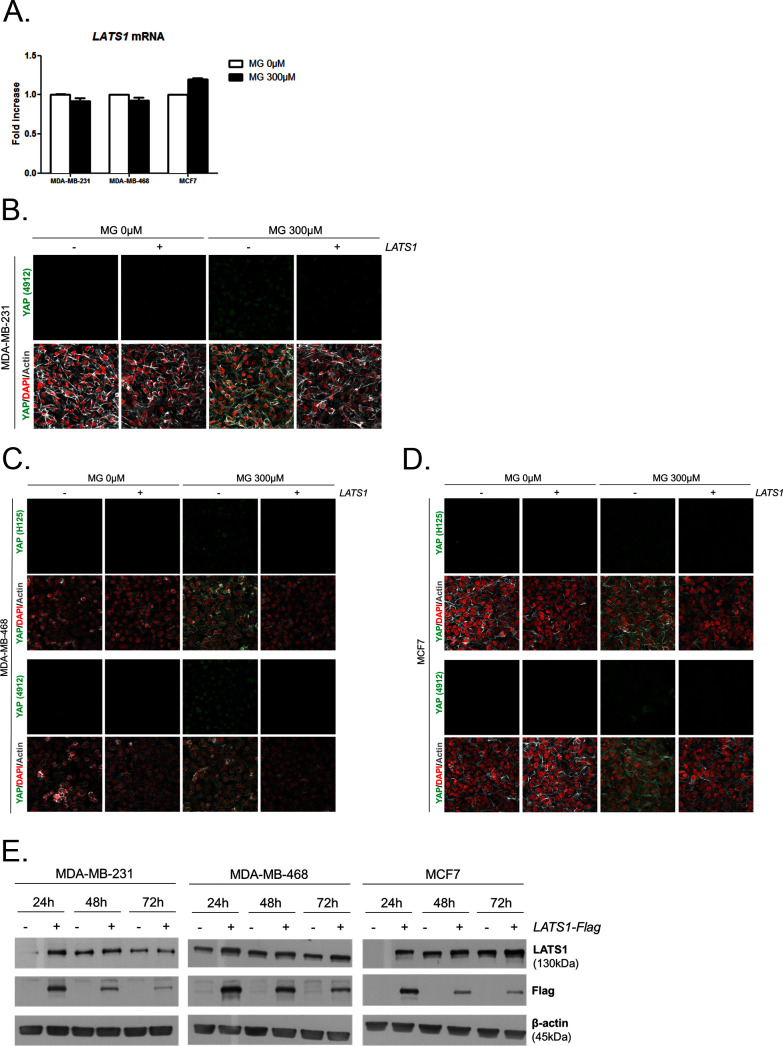
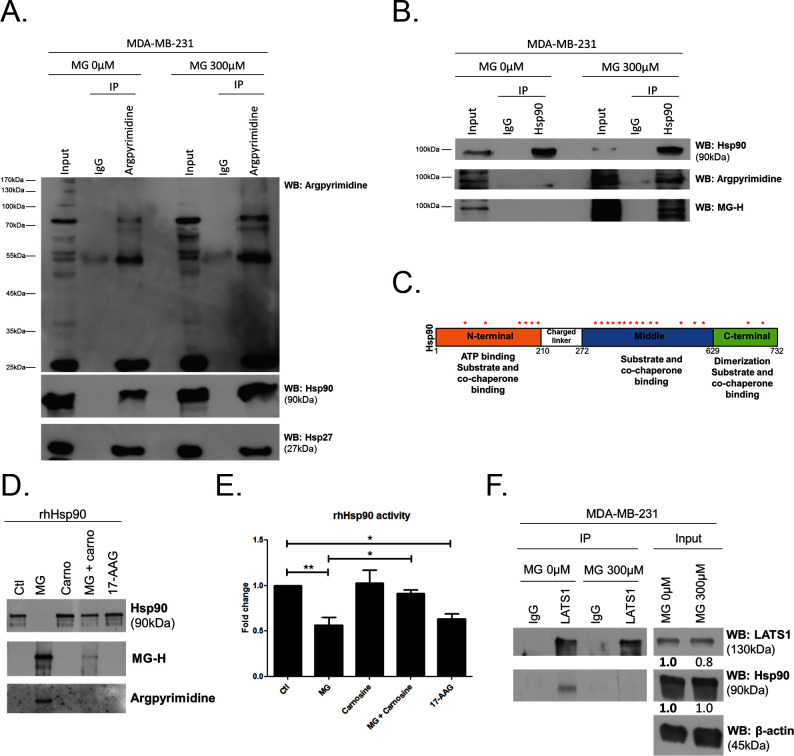
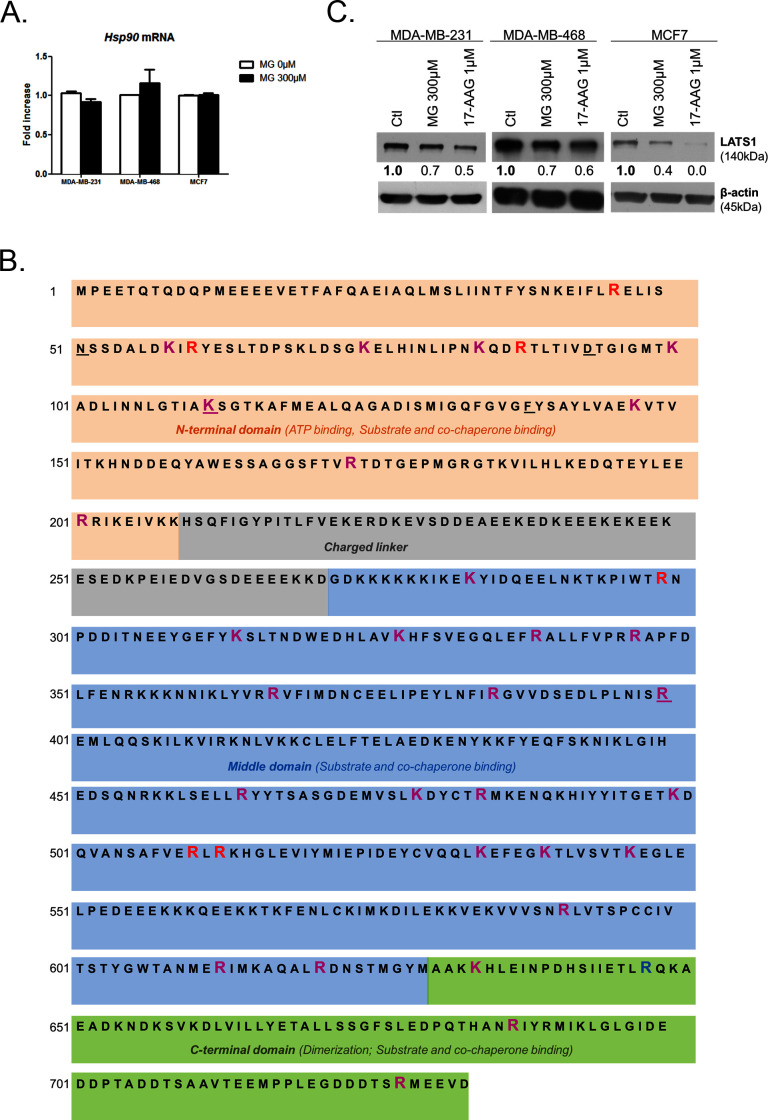
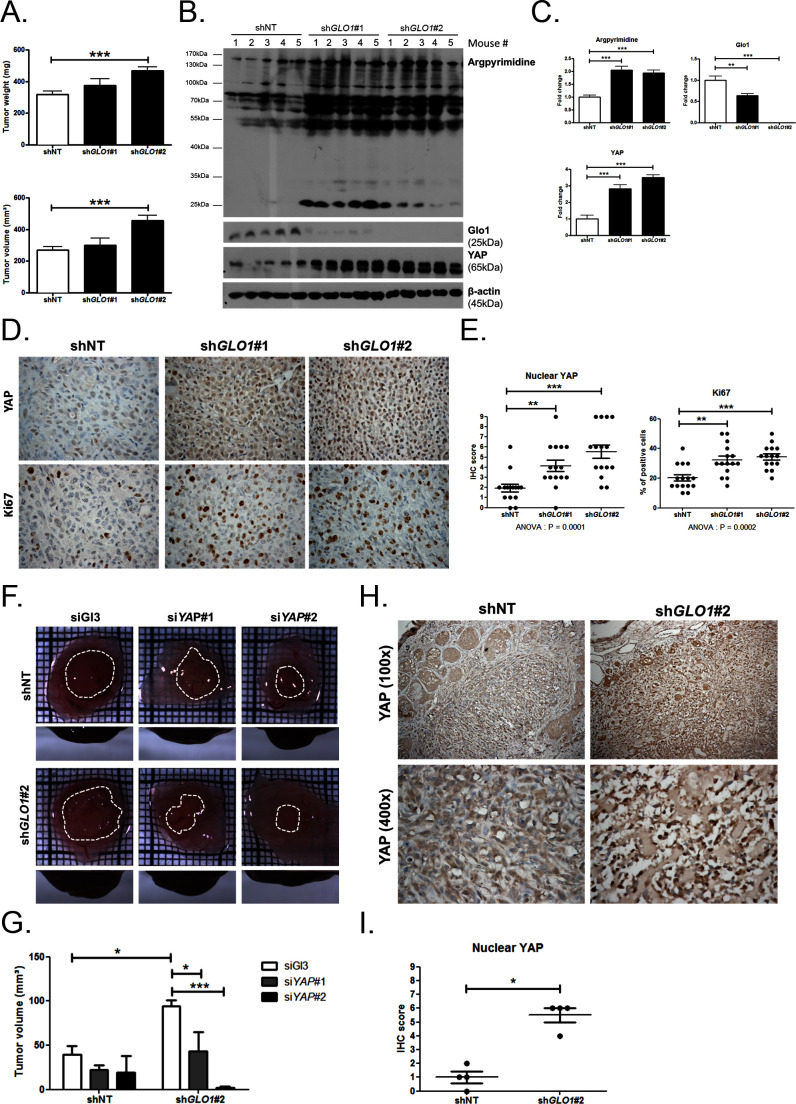
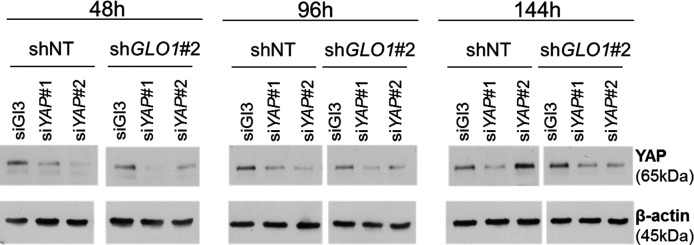
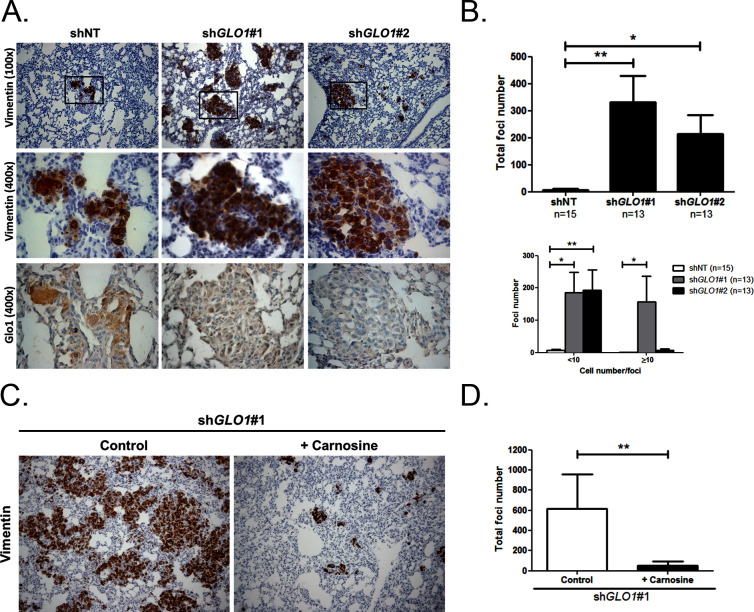
Metabolic reprogramming toward aerobic glycolysis unavoidably induces methylglyoxal (MG) formation in cancer cells. MG mediates the glycation of proteins to form advanced glycation end products (AGEs). We have recently demonstrated that MG-induced AGEs are a common feature of breast cancer. Little is known regarding the impact of MG-mediated carbonyl stress on tumor progression. Breast tumors with MG stress presented with high nuclear YAP, a key transcriptional co-activator regulating tumor growth and invasion. Elevated MG levels resulted in sustained YAP nuclear localization/activity that could be reverted using Carnosine, a scavenger for MG. MG treatment affected Hsp90 chaperone activity and decreased its binding to LATS1, a key kinase of the Hippo pathway. Cancer cells with high MG stress showed enhanced growth and metastatic potential in vivo. These findings reinforce the cumulative evidence pointing to hyperglycemia as a risk factor for cancer incidence and bring renewed interest in MG scavengers for cancer treatment.
癌细胞向有氧糖酵解的代谢重编程不可避免地会诱导甲基乙二醛(MG)的形成。MG介导蛋白质糖基化形成晚期糖基化终产物(AGEs)。我们最近证明,MG诱导的AGEs是乳腺癌的一个常见特征。关于MG介导的羰基应激对肿瘤进展的影响知之甚少。存在MG应激的乳腺肿瘤表现为高核YAP,YAP是一种调节肿瘤生长和侵袭的关键转录共激活因子。MG水平升高导致YAP持续核定位/活性,使用MG清除剂肌肽可使其恢复。MG处理影响Hsp90伴侣活性,并降低其与Hippo通路关键激酶LATS1的结合。具有高MG应激的癌细胞在体内显示出增强的生长和转移潜能。这些发现强化了越来越多的证据,表明高血糖是癌症发生的危险因素,并重新引发了人们对MG清除剂用于癌症治疗的兴趣。