Department of Biochemistry and Molecular Biology, The Pennsylvania State University, University Park, Pennsylvania, United States of America.
Huck Institutes of the Life Sciences, The Pennsylvania State University, University Park, Pennsylvania, United States of America.
PLoS Pathog. 2018 Apr 27;14(4):e1007036. doi: 10.1371/journal.ppat.1007036. eCollection 2018 Apr.
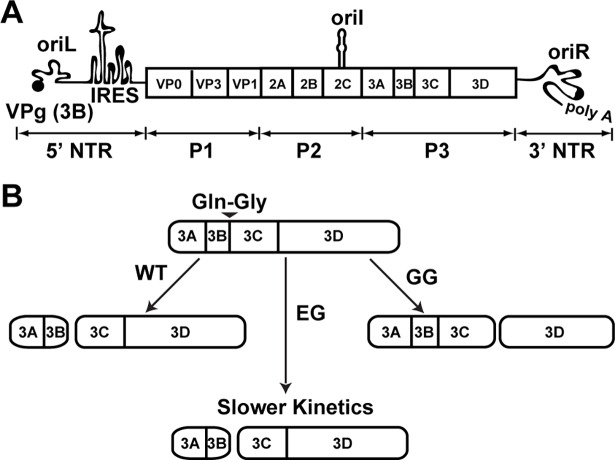
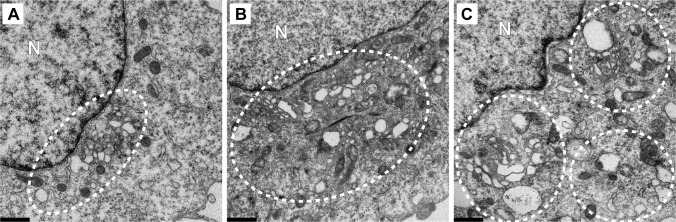
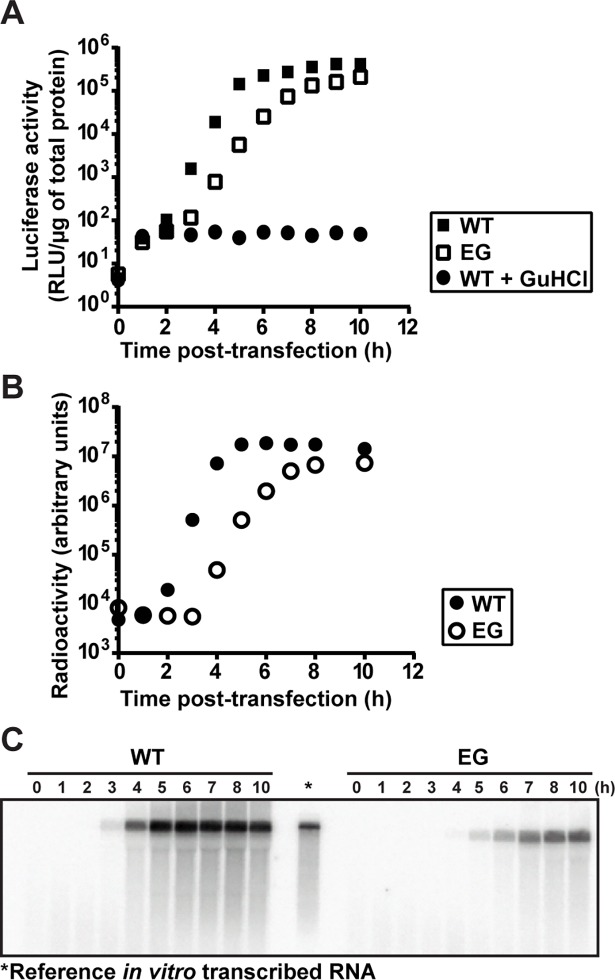
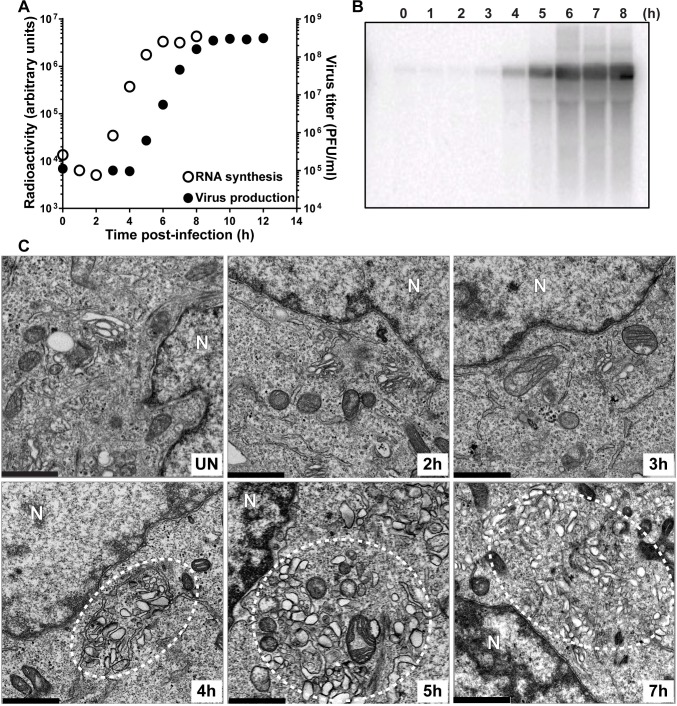
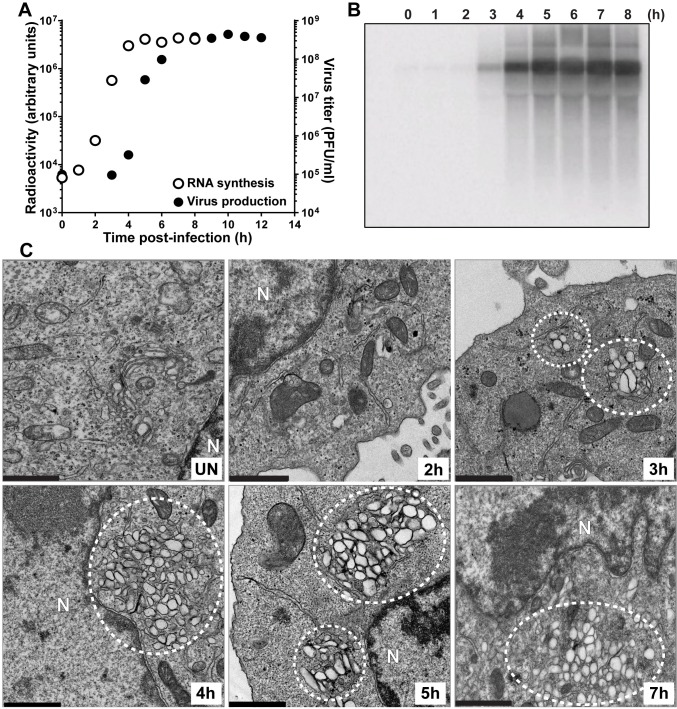
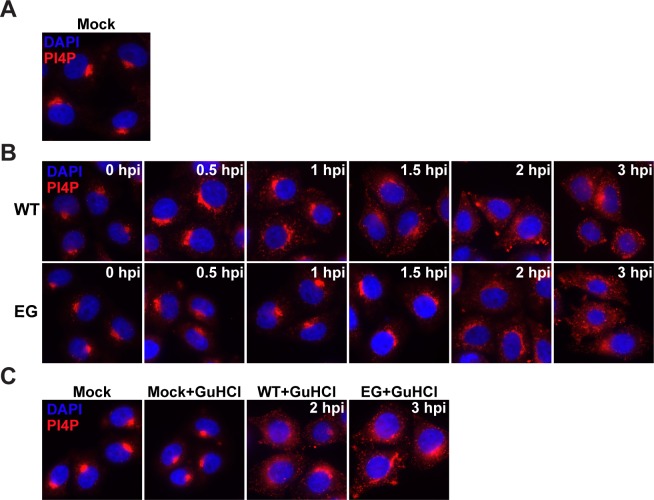
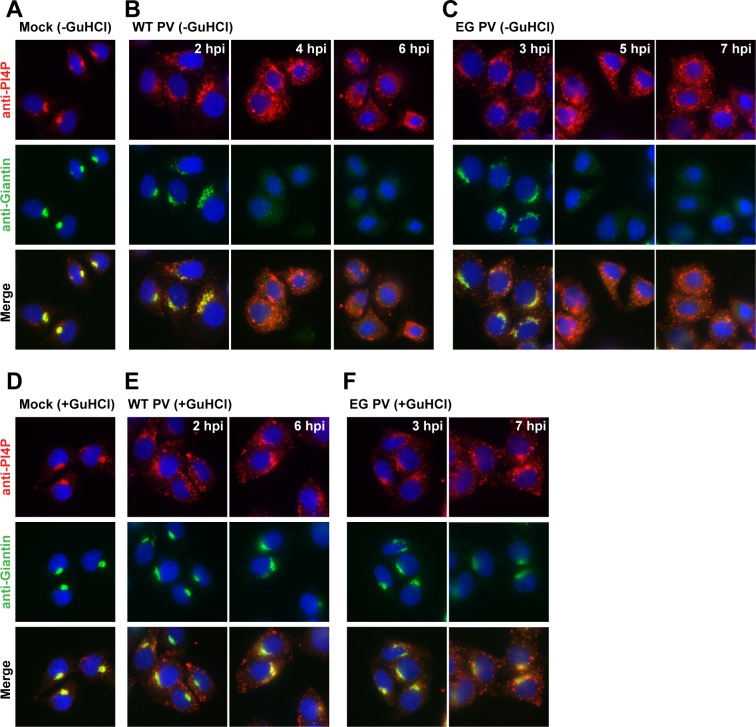
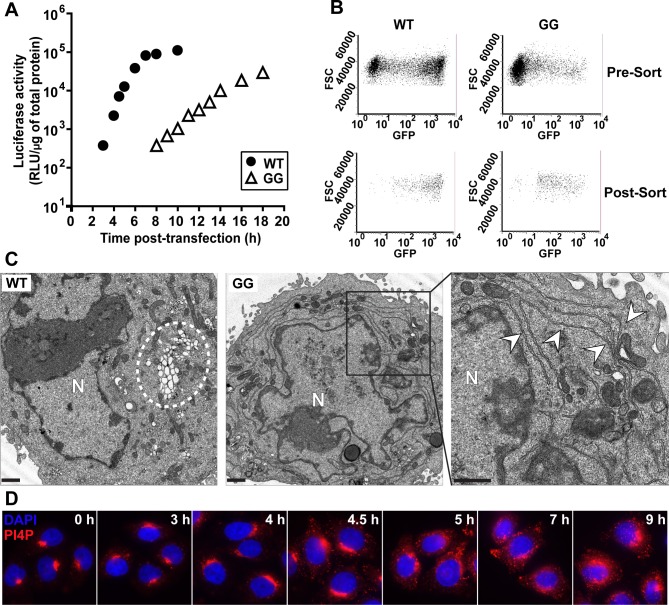
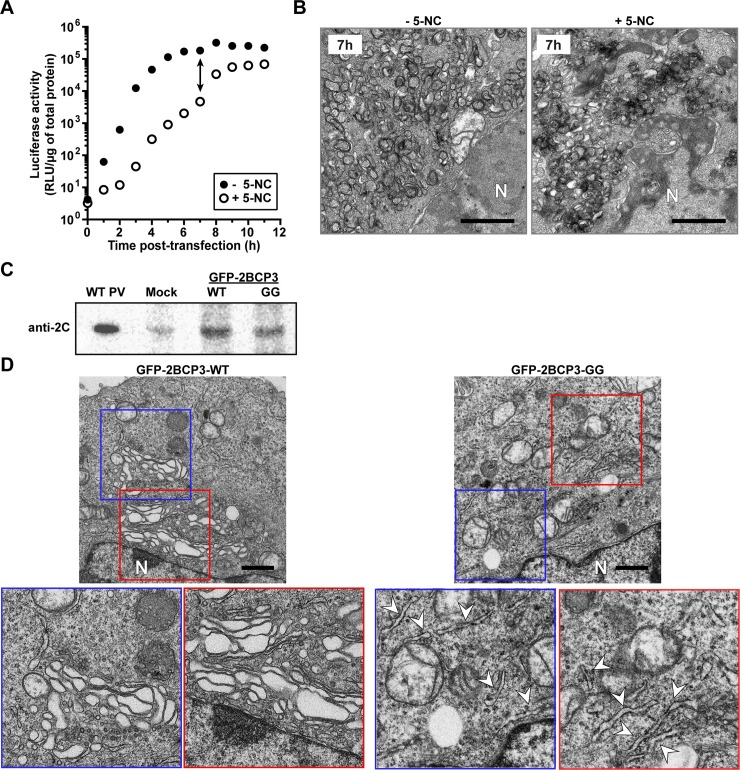
At the culmination of poliovirus (PV) multiplication, membranes are observed that contain phosphatidylinositol-4-phosphate (PI4P) and appear as vesicular clusters in cross section. Induction and remodeling of PI4P and membranes prior to or concurrent with genome replication has not been well studied. Here, we exploit two PV mutants, termed EG and GG, which exhibit aberrant proteolytic processing of the P3 precursor that substantially delays the onset of genome replication and/or impairs virus assembly, to illuminate the pathway of formation of PV-induced membranous structures. For WT PV, changes to the PI4P pool were observed as early as 30 min post-infection. PI4P remodeling occurred even in the presence of guanidine hydrochloride, a replication inhibitor, and was accompanied by formation of membrane tubules throughout the cytoplasm. Vesicular clusters appeared in the perinuclear region of the cell at 3 h post-infection, a time too slow for these structures to be responsible for genome replication. Delays in the onset of genome replication observed for EG and GG PVs were similar to the delays in virus-induced remodeling of PI4P pools, consistent with PI4P serving as a marker of the genome-replication organelle. GG PV was unable to convert virus-induced tubules into vesicular clusters, perhaps explaining the nearly 5-log reduction in infectious virus produced by this mutant. Our results are consistent with PV inducing temporally distinct membranous structures (organelles) for genome replication (tubules) and virus assembly (vesicular clusters). We suggest that the pace of formation, spatiotemporal dynamics, and the efficiency of the replication-to-assembly-organelle conversion may be set by both the rate of P3 polyprotein processing and the capacity for P3 processing to yield 3AB and/or 3CD proteins.
在脊髓灰质炎病毒(PV)复制的末期,可以观察到含有磷脂酰肌醇-4-磷酸(PI4P)的膜,这些膜在横截面上呈现出囊泡簇的形式。在基因组复制之前或同时,PI4P 和膜的诱导和重塑尚未得到很好的研究。在这里,我们利用两种 PV 突变体,称为 EG 和 GG,它们表现出 P3 前体异常的蛋白水解加工,这大大延迟了基因组复制的开始和/或损害了病毒组装,以阐明 PV 诱导的膜结构形成途径。对于 WT PV,在感染后 30 分钟即可观察到 PI4P 池的变化。PI4P 的重塑甚至在胍盐酸盐(一种复制抑制剂)存在的情况下也会发生,并且伴随着整个细胞质中膜小管的形成。在感染后 3 小时,囊泡簇出现在核周区域,这个时间对于这些结构负责基因组复制来说太慢了。EG 和 GG PV 中观察到的基因组复制开始延迟与 PI4P 池病毒诱导重塑的延迟相似,这与 PI4P 作为基因组复制细胞器的标志物一致。GG PV 无法将病毒诱导的小管转化为囊泡簇,这也许可以解释这种突变体产生的感染性病毒减少近 5 个对数级。我们的结果与 PV 诱导用于基因组复制(小管)和病毒组装(囊泡簇)的时间上不同的膜结构(细胞器)一致。我们认为形成的速度、时空动态以及复制到组装细胞器的转换效率可能由 P3 多蛋白加工的速度和 P3 加工产生 3AB 和/或 3CD 蛋白的能力决定。