Department of Chemistry and Biochemistry , Utah State University , Logan , Utah 84322 , United States.
Pacific Northwest National Laboratory , Richland , Washington 99352 , United States.
Acc Chem Res. 2018 Sep 18;51(9):2179-2186. doi: 10.1021/acs.accounts.8b00112. Epub 2018 Aug 10.
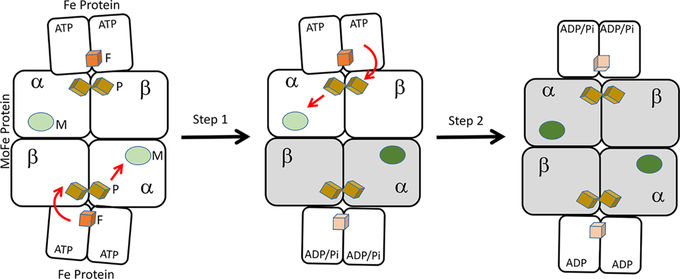
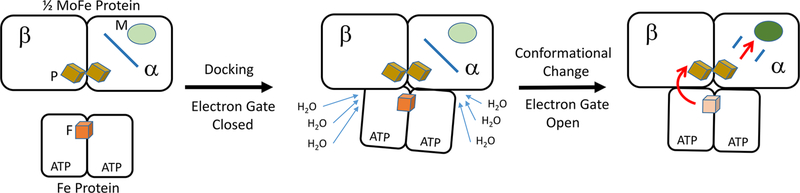
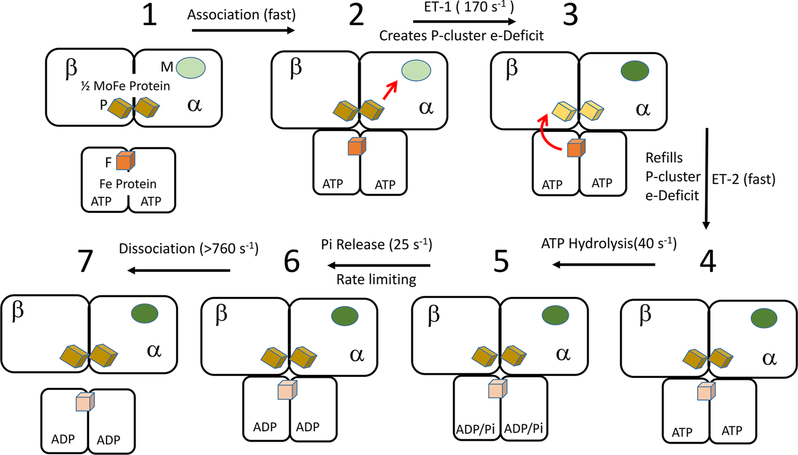
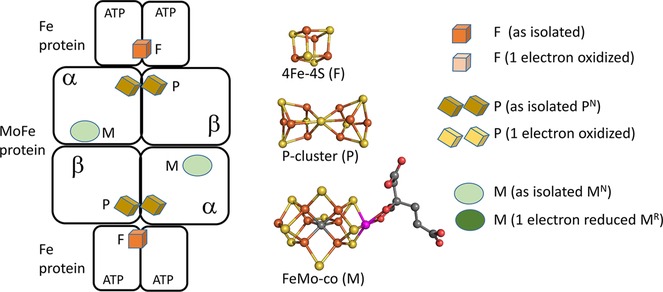
Nitrogenase is a complicated two-component enzyme system that uses ATP binding and hydrolysis energy to achieve one of the most difficult chemical reactions in nature, the reduction of N to NH. One component of the Mo-based nitrogenase system, Fe protein, delivers electrons one at a time to the second component, the catalytic MoFe protein. This process occurs through a series of synchronized events collectively called the "Fe protein cycle". Elucidating details of the events associated with this cycle has constituted an important challenge in understanding the nitrogenase mechanism. Electron delivery is a multistep process involving three metal clusters with intra- and interprotein events. It is proposed that the first electron transfer event is a gated intraprotein transfer of one electron from the MoFe protein P-cluster to the FeMo cofactor. Measurement of the effect of osmotic pressure on the rate of this electron transfer process revealed that it is gated by protein conformational changes. This first electron transfer is activated by binding of the Fe protein containing two bound ATP molecules. The mechanism of how this protein-protein association triggers electron transfer remains unknown. The second electron transfer event is proposed to be a rapid interprotein "backfill" with transfer of one electron from the reduced Fe protein 4Fe-4S cluster to the oxidized P-cluster. In this way, electron delivery can be viewed as a case of "deficit spending". Such a deficit-spending electron transfer process can be envisioned as a way to achieve one-direction electron flow, limiting the potential for back electron flow. Hydrolysis of two ATP molecules associated with the Fe protein occurs after the electron transfer and therefore is not used to directly drive the electron transfer. Rather, ATP hydrolysis is proposed to contribute to relaxation of the "activated" conformational state associated with the ATP form of the complex, with the free energy from ATP hydrolysis being used to pay back energy associated with component protein association and electron transfer. Release of inorganic phosphate (Pi) and protein-protein dissociation follow electron transfer and ATP hydrolysis. The rate-limiting step for the Fe protein cycle is not dissociation of the two proteins, as previously believed, but rather is release of Pi after ATP hydrolysis, which is then followed by rapid protein-protein complex dissociation. Nitrogenase is composed of two catalytic halves that do not function independently but rather exhibit anticooperative nuclear motion in which electron transfer in one-half of the complex partially inhibits electron transfer and ATP hydrolysis in the other half. Calculations indicated the existence of anticooperative interactions across the entire nitrogenase complex, suggesting a mechanism for the control of events on opposite ends of this large complex. The mechanistic necessity for this anticooperative process remains unknown. This Account presents a working model for how all of these processes work together in the nitrogenase "machine" to transduce the energy from ATP binding and hydrolysis to drive N reduction.
固氮酶是一种复杂的双组分酶系统,它利用 ATP 结合和水解能量来实现自然界中最困难的化学反应之一,即 N 到 NH 的还原。钼基固氮酶系统的一个组成部分,铁蛋白,一次将电子传递给第二个组成部分,即催化的 MoFe 蛋白。这个过程通过一系列称为“铁蛋白循环”的同步事件发生。阐明与该循环相关的事件细节一直是理解固氮酶机制的重要挑战。电子传递是一个多步骤过程,涉及三个带有蛋白内和蛋白间事件的金属簇。据推测,第一个电子转移事件是 MoFe 蛋白 P 簇中的一个电子从 MoFe 蛋白 P 簇到 FeMo 辅因子的门控蛋白内转移。测量渗透压对这个电子转移过程速率的影响表明,它是由蛋白质构象变化门控的。这个第一个电子转移被结合了两个结合的 ATP 分子的铁蛋白激活。这个蛋白质-蛋白质缔合如何触发电子转移的机制仍然未知。第二个电子转移事件被提议为一个快速的蛋白间“回填”,其中一个电子从还原的 Fe 蛋白 4Fe-4S 簇转移到氧化的 P 簇。这样,电子传递可以被视为“赤字支出”的情况。这种赤字支出的电子传递过程可以被设想为实现单向电子流的一种方式,限制了反向电子流的潜力。与铁蛋白相关的两个 ATP 分子的水解发生在电子转移之后,因此不能直接驱动电子转移。相反,据推测,ATP 水解有助于与复合物的 ATP 形式相关的“激活”构象状态的松弛,ATP 水解产生的自由能用于偿还与组件蛋白缔合和电子转移相关的能量。无机磷酸盐 (Pi) 的释放和蛋白质-蛋白质的解离紧随电子转移和 ATP 水解之后。铁蛋白循环的限速步骤不是两种蛋白质的解离,如以前所认为的那样,而是 ATP 水解后的 Pi 释放,然后是快速的蛋白质-蛋白质复合物解离。固氮酶由两个催化半体组成,它们不能独立发挥作用,而是表现出反协同核运动,其中复合物一半的电子转移部分抑制另一半的电子转移和 ATP 水解。计算表明,整个固氮酶复合物存在反协同相互作用,这表明了一种控制这个大型复合物两端事件的机制。这种反协同过程的机械必要性仍然未知。本账户介绍了一个工作模型,说明这些过程如何共同作用于固氮酶“机器”,将 ATP 结合和水解的能量转化为 N 还原的驱动力。