Center for Models of Life, Niels Bohr Institute, University of Copenhagen, Copenhagen, Denmark.
StemPhys, Niels Bohr Institute, University of Copenhagen, Copenhagen, Denmark.
Elife. 2018 Nov 27;7:e38407. doi: 10.7554/eLife.38407.
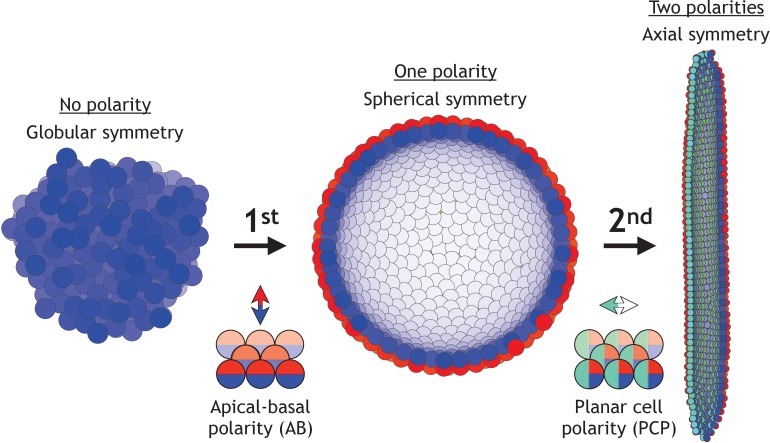
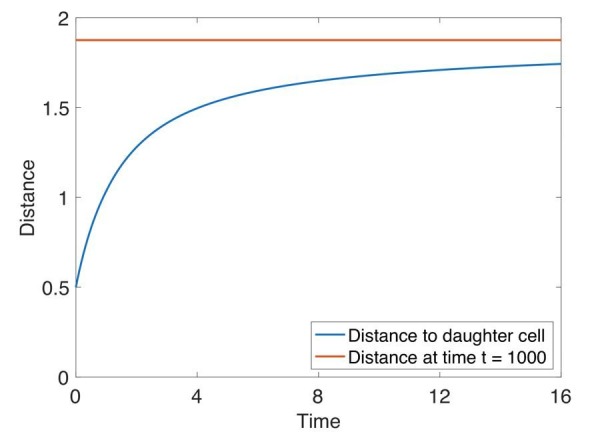
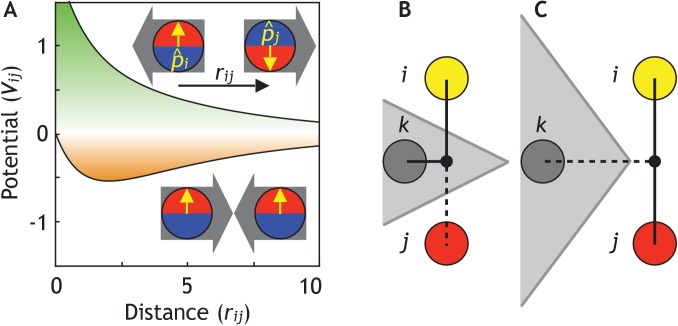
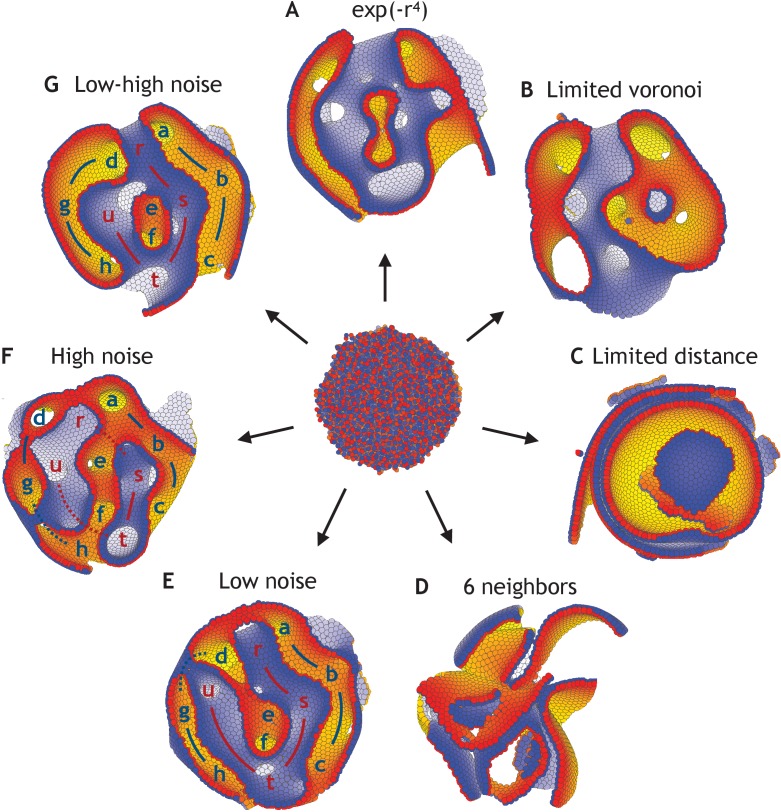
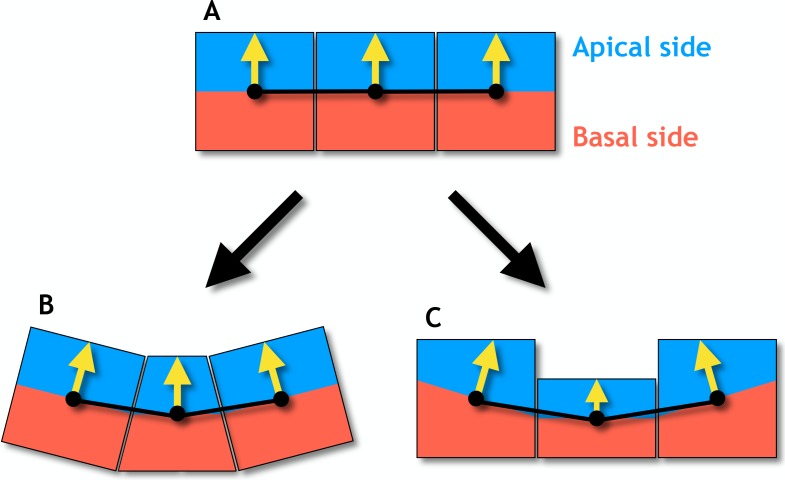
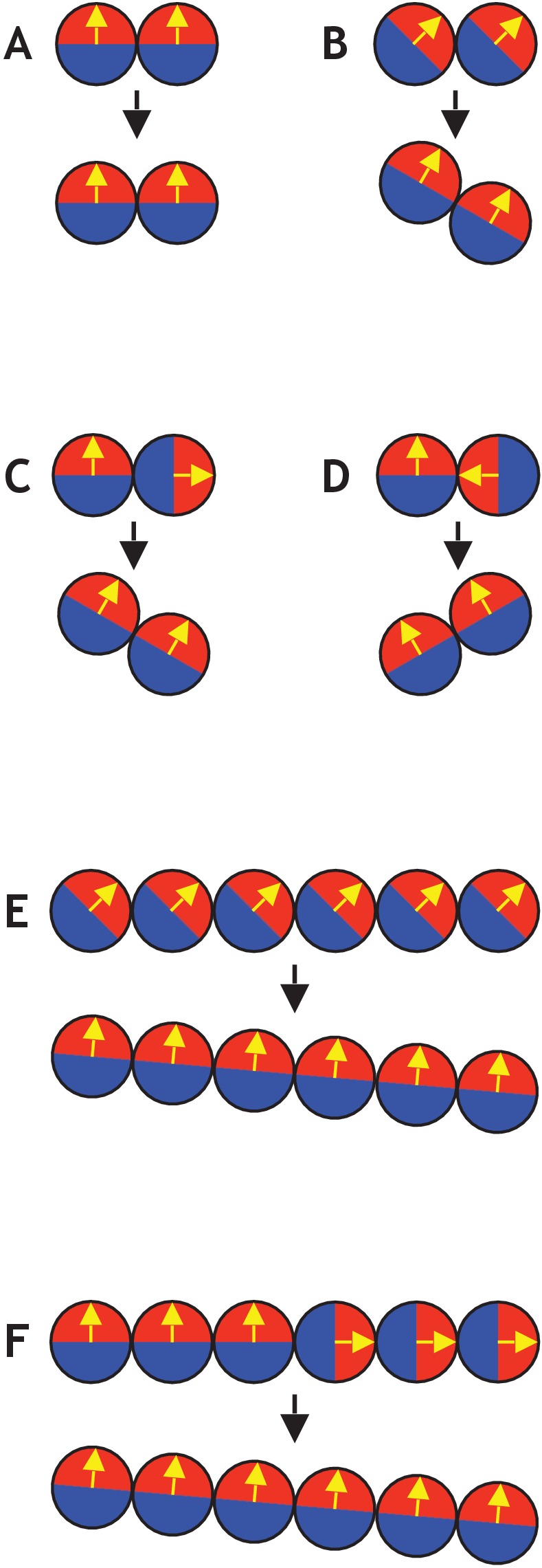
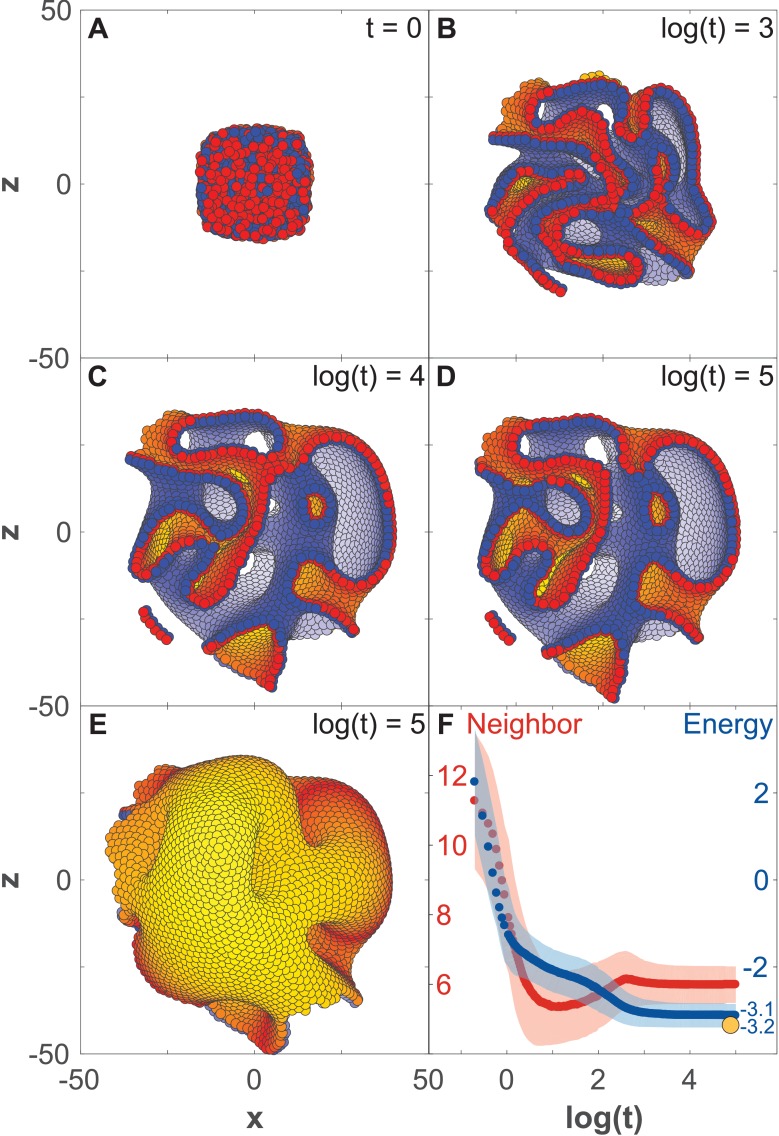
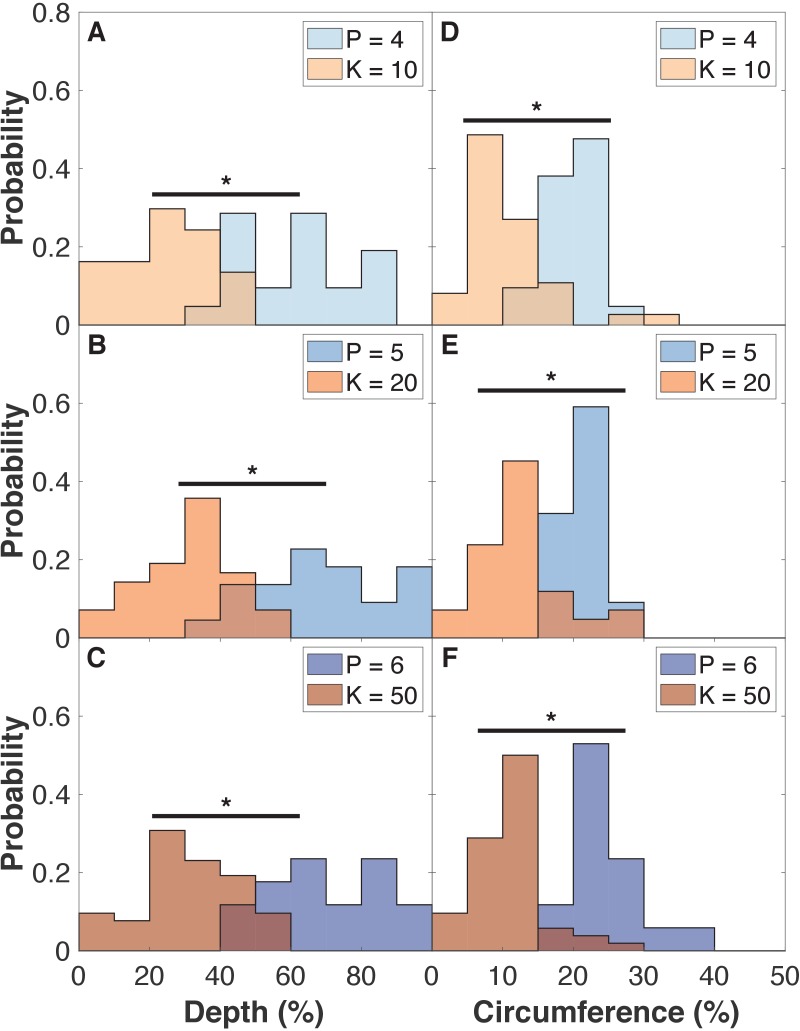
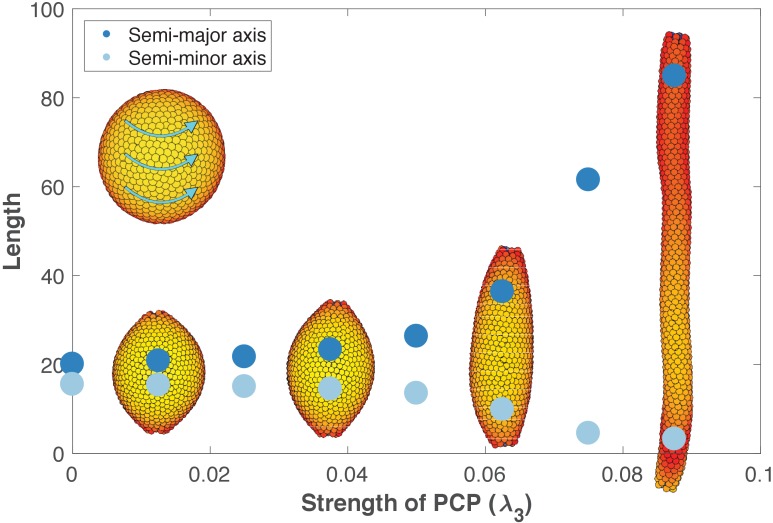
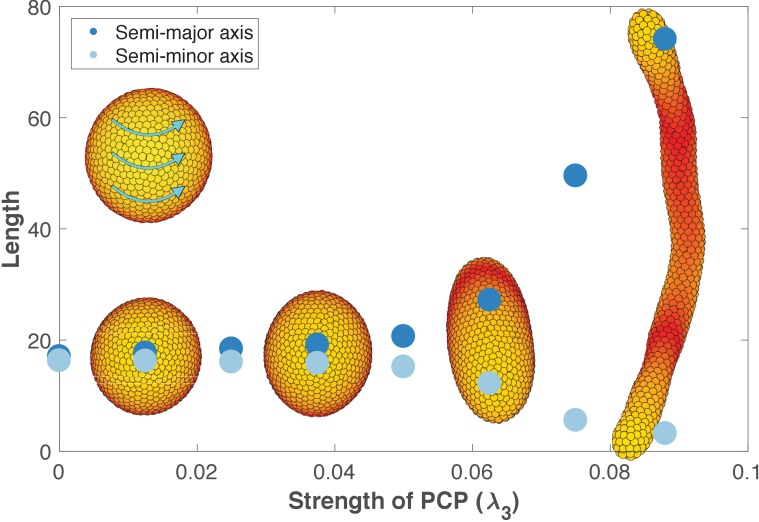
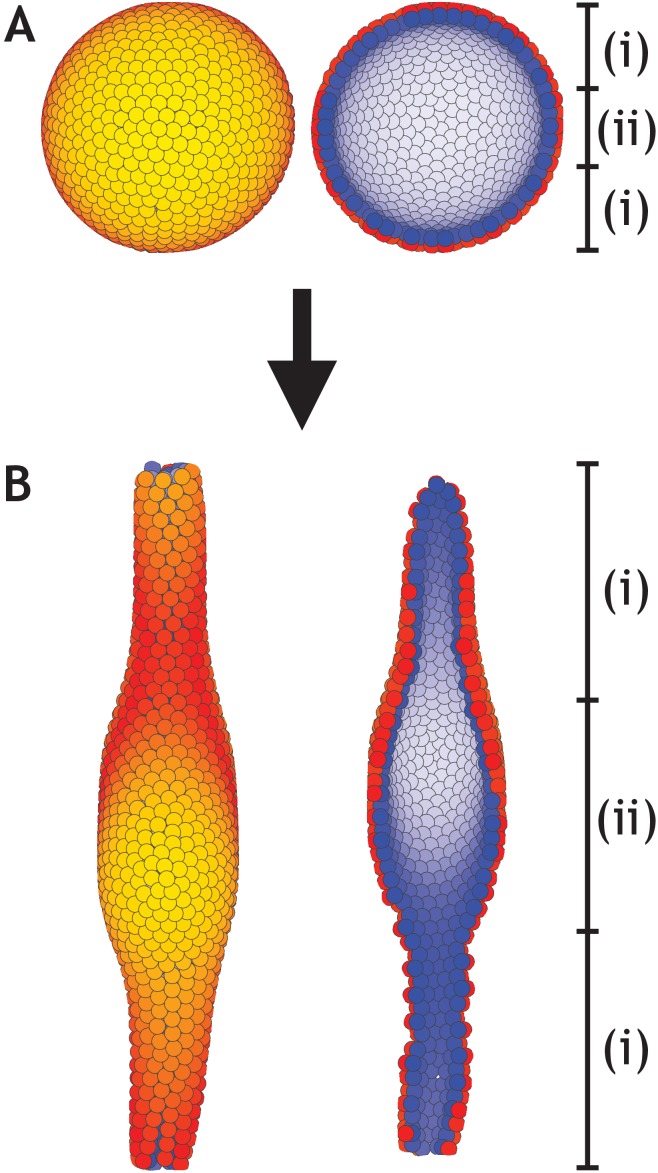
Despite continual renewal and damages, a multicellular organism is able to maintain its complex morphology. How is this stability compatible with the complexity and diversity of living forms? Looking for answers at protein level may be limiting as diverging protein sequences can result in similar morphologies. Inspired by the progressive role of apical-basal and planar cell polarity in development, we propose that stability, complexity, and diversity are emergent properties in populations of proliferating polarized cells. We support our hypothesis by a theoretical approach, developed to effectively capture both types of polar cell adhesions. When applied to specific cases of development - gastrulation and the origins of folds and tubes - our theoretical tool suggests experimentally testable predictions pointing to the strength of polar adhesion, restricted directions of cell polarities, and the rate of cell proliferation to be major determinants of morphological diversity and stability.
尽管不断更新和损坏,多细胞生物仍能够维持其复杂的形态。这种稳定性如何与生物形态的复杂性和多样性相兼容?在蛋白质水平上寻找答案可能具有局限性,因为不同的蛋白质序列可能导致相似的形态。受顶端-基底和平面细胞极性在发育中渐进作用的启发,我们提出稳定性、复杂性和多样性是增殖极化细胞群体中的涌现属性。我们通过一种理论方法来支持我们的假设,该方法旨在有效地捕捉两种类型的极性细胞粘附。当应用于发育的具体情况 - 原肠胚形成和褶皱和管的起源 - 我们的理论工具表明了可进行实验验证的预测,指出了极性粘附的强度、细胞极性的受限方向以及细胞增殖率是形态多样性和稳定性的主要决定因素。