Wang Yundi, Eldstrom Jodene, Fedida David
Department of Anesthesiology, Pharmacology & Therapeutics, The University of British Columbia, Vancouver, BC, Canada.
Front Physiol. 2020 Jun 4;11:504. doi: 10.3389/fphys.2020.00504. eCollection 2020.
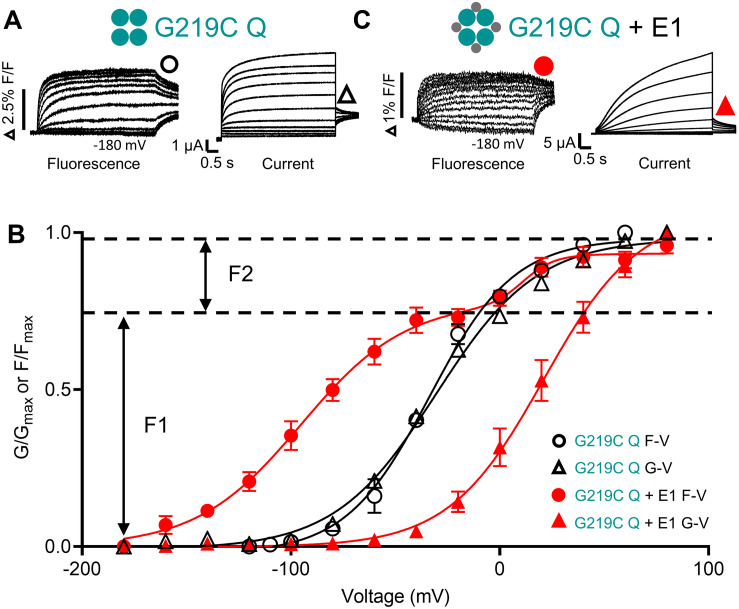
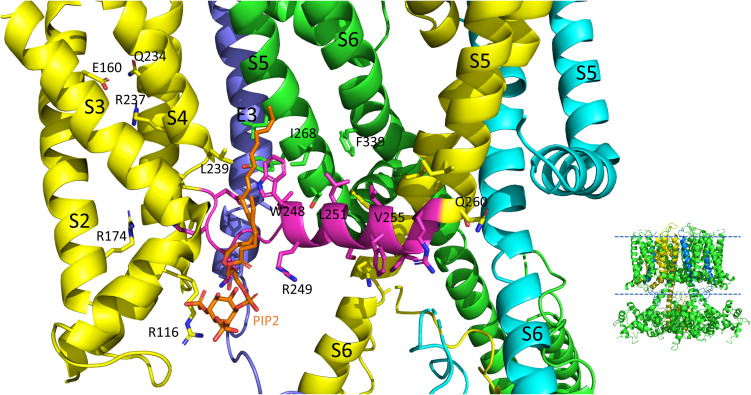
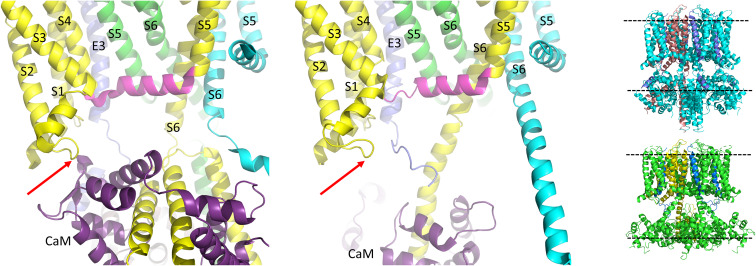
The IKs channel complex is formed by the co-assembly of Kv7.1 (KCNQ1), a voltage-gated potassium channel, with its β-subunit, KCNE1 and the association of numerous accessory regulatory molecules such as PIP2, calmodulin, and yotiao. As a result, the IKs potassium current shows kinetic and regulatory flexibility, which not only allows IKs to fulfill physiological roles as disparate as cardiac repolarization and the maintenance of endolymph K homeostasis, but also to cause significant disease when it malfunctions. Here, we review new areas of understanding in the assembly, kinetics of activation and inactivation, voltage-sensor pore coupling, unitary events and regulation of this important ion channel complex, all of which have been given further impetus by the recent solution of cryo-EM structural representations of KCNQ1 alone and KCNQ1+KCNE3. Recently, the stoichiometric ratio of KCNE1 to KCNQ1 subunits has been confirmed to be variable up to a ratio of 4:4, rather than fixed at 2:4, and we will review the results and new methodologies that support this conclusion. Significant advances have been made in understanding differences between KCNQ1 and IKs gating using voltage clamp fluorimetry and mutational analysis to illuminate voltage sensor activation and inactivation, and the relationship between voltage sensor translation and pore domain opening. We now understand that the KCNQ1 pore can open with different permeabilities and conductance when the voltage sensor is in partially or fully activated positions, and the ability to make robust single channel recordings from IKs channels has also revealed the complicated pore subconductance architecture during these opening steps, during inactivation, and regulation by 1-4 associated KCNE1 subunits. Experiments placing mutations into individual voltage sensors to drastically change voltage dependence or prevent their movement altogether have demonstrated that the activation of KCNQ1 alone and IKs can best be explained using allosteric models of channel gating. Finally, we discuss how the intrinsic gating properties of KCNQ1 and IKs are highly modulated through the impact of intracellular signaling molecules and co-factors such as PIP2, protein kinase A, calmodulin and ATP, all of which modulate IKs current kinetics and contribute to diverse IKs channel complex function.
IKs通道复合体由电压门控钾通道Kv7.1(KCNQ1)与其β亚基KCNE1共同组装而成,并与众多辅助调节分子如磷脂酰肌醇-4,5-二磷酸(PIP2)、钙调蛋白和Yotiao相关联。因此,IKs钾电流表现出动力学和调节的灵活性,这不仅使IKs能够发挥从心脏复极化到维持内淋巴钾离子稳态等截然不同的生理作用,而且当其功能异常时还会引发重大疾病。在此,我们综述了对这个重要离子通道复合体的组装、激活和失活动力学、电压传感器-孔道偶联、单一事件及调节等方面的新认识领域,最近单独的KCNQ1以及KCNQ1+KCNE3的冷冻电镜结构表征的解析进一步推动了这些研究。最近,已证实KCNE1与KCNQ1亚基的化学计量比在4:4范围内可变,而非固定为2:4,我们将综述支持这一结论的研究结果和新方法。利用电压钳荧光法和突变分析来阐明电压传感器的激活和失活以及电压传感器平移与孔道结构域开放之间的关系,在理解KCNQ1和IKs门控差异方面取得了重大进展。我们现在明白,当电压传感器处于部分或完全激活位置时,KCNQ1孔道可以不同的通透性和电导率开放,并且能够从IKs通道进行可靠的单通道记录也揭示了这些开放步骤、失活过程以及1-4个相关KCNE1亚基调节过程中复杂的孔道亚电导结构。将突变引入单个电压传感器以大幅改变电压依赖性或完全阻止其移动的实验表明,使用通道门控的变构模型可以最好地解释单独的KCNQ1和IKs的激活。最后,我们讨论KCNQ1和IKs的内在门控特性如何通过细胞内信号分子和辅助因子如PIP2、蛋白激酶A、钙调蛋白和ATP的影响而受到高度调节,所有这些都调节IKs电流动力学并有助于IKs通道复合体发挥多种功能。