Gladstone Institutes, San Francisco, CA, 94158, USA.
Roddenberry Center for Stem Cell Biology and Medicine at Gladstone, San Francisco, CA, 94158, USA.
Nat Commun. 2020 Nov 5;11(1):5612. doi: 10.1038/s41467-020-19283-x.
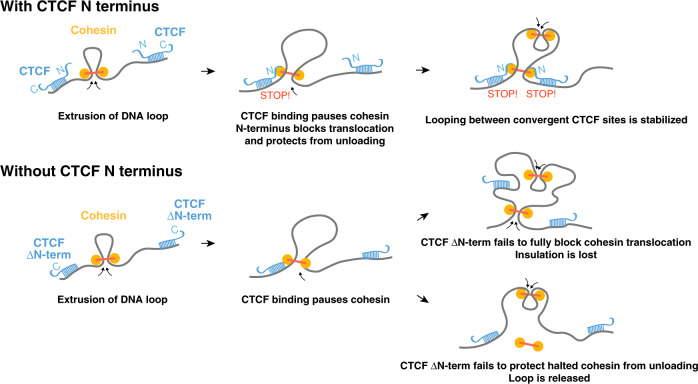
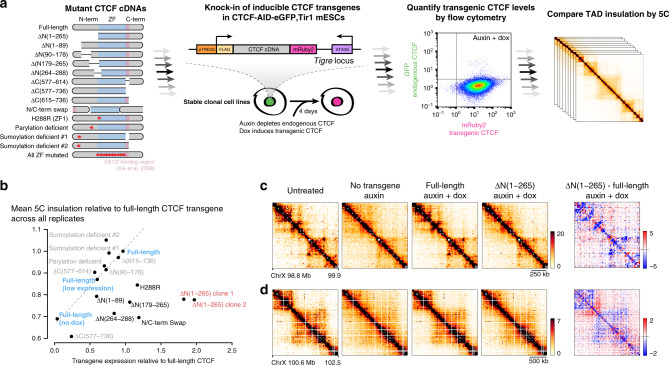
Current models propose that boundaries of mammalian topologically associating domains (TADs) arise from the ability of the CTCF protein to stop extrusion of chromatin loops by cohesin. While the orientation of CTCF motifs determines which pairs of CTCF sites preferentially stabilize loops, the molecular basis of this polarity remains unclear. By combining ChIP-seq and single molecule live imaging we report that CTCF positions cohesin, but does not control its overall binding dynamics on chromatin. Using an inducible complementation system, we find that CTCF mutants lacking the N-terminus cannot insulate TADs properly. Cohesin remains at CTCF sites in this mutant, albeit with reduced enrichment. Given the orientation of CTCF motifs presents the N-terminus towards cohesin as it translocates from the interior of TADs, these observations explain how the orientation of CTCF binding sites translates into genome folding patterns.
目前的模型提出,哺乳动物拓扑关联域 (TAD) 的边界源于 CTCF 蛋白阻止黏连蛋白挤出染色质环的能力。虽然 CTCF 基序的取向决定了哪些对 CTCF 位点优先稳定环,但这种极性的分子基础仍不清楚。通过结合 ChIP-seq 和单分子实时成像,我们报告 CTCF 定位黏连蛋白,但不控制其在染色质上的整体结合动力学。使用诱导互补系统,我们发现缺乏 N 端的 CTCF 突变体不能正确隔离 TAD。尽管在这种突变体中,黏连蛋白的富集减少了,但它仍然存在于 CTCF 位点。鉴于 CTCF 基序的取向在 CTCF 从 TAD 内部易位时将 N 端朝向黏连蛋白,这些观察结果解释了 CTCF 结合位点的取向如何转化为基因组折叠模式。