Department of Biochemistry and Molecular Biology, Michigan State University, East Lansing, United States.
Department of Physics, Michigan State University, East Lansing, United States.
Elife. 2021 Jan 26;10:e64004. doi: 10.7554/eLife.64004.
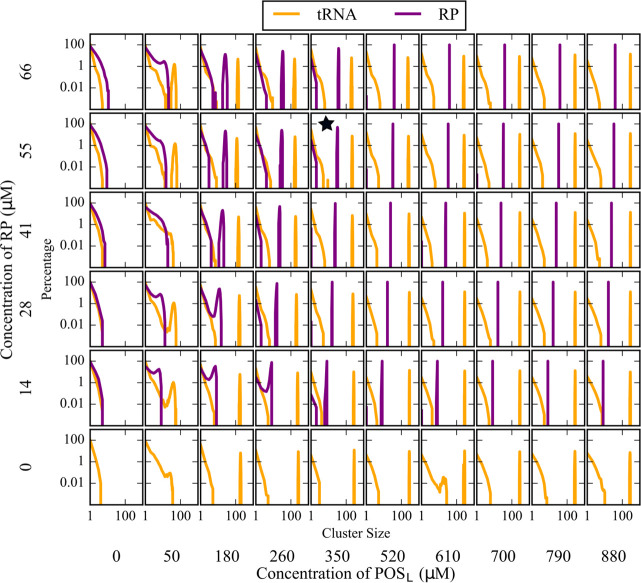
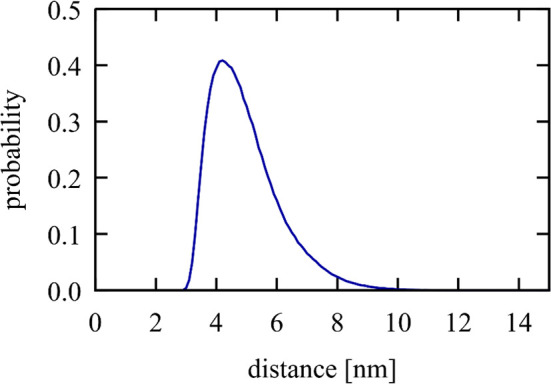
Phase separation processes are increasingly being recognized as important organizing mechanisms of biological macromolecules in cellular environments. Well-established drivers of phase separation are multi-valency and intrinsic disorder. Here, we show that globular macromolecules may condense simply based on electrostatic complementarity. More specifically, phase separation of mixtures between RNA and positively charged proteins is described from a combination of multiscale computer simulations with microscopy and spectroscopy experiments. Phase diagrams were mapped out as a function of molecular concentrations in experiment and as a function of molecular size and temperature via simulations. The resulting condensates were found to retain at least some degree of internal dynamics varying as a function of the molecular composition. The results suggest a more general principle for phase separation that is based primarily on electrostatic complementarity without invoking polymer properties as in most previous studies. Simulation results furthermore suggest that such phase separation may occur widely in heterogenous cellular environment between nucleic acid and protein components.
相分离过程越来越被认为是细胞环境中生物大分子的重要组织机制。相分离的既定驱动因素是多价和固有无序。在这里,我们表明,球形大分子可能仅仅基于静电互补而凝聚。更具体地说,我们通过多尺度计算机模拟与显微镜和光谱实验的组合,描述了 RNA 和带正电荷的蛋白质之间混合物的相分离。通过实验绘制了相图,作为分子浓度的函数,并通过模拟作为分子大小和温度的函数绘制了相图。结果发现,所得的凝聚物至少保留了一定程度的内部动力学,该动力学随分子组成的变化而变化。结果表明,相分离的一个更普遍的原则主要基于静电互补,而不像大多数先前的研究那样,援引聚合物性质。模拟结果还表明,这种相分离可能在核酸和蛋白质成分之间的异质细胞环境中广泛发生。