Department of Cellular and Molecular Pharmacology, University of California at San Francisco, San Francisco, California, United States.
Department of Neurobiology, Stanford University School of Medicine, Stanford, California, United States.
Physiol Rev. 2023 Oct 1;103(4):2877-2925. doi: 10.1152/physrev.00034.2022. Epub 2023 Jun 8.
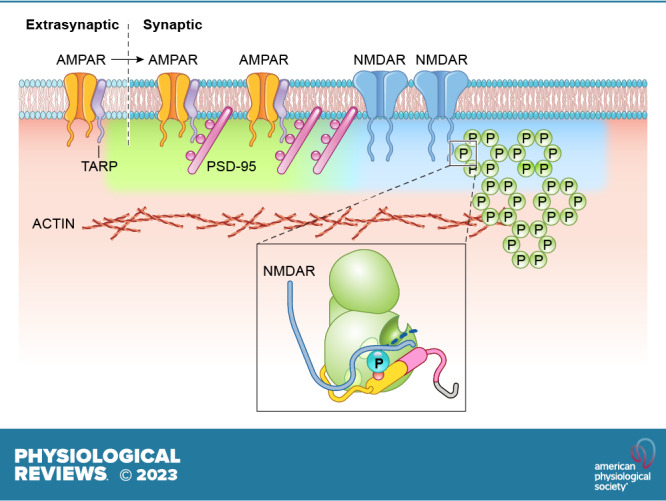
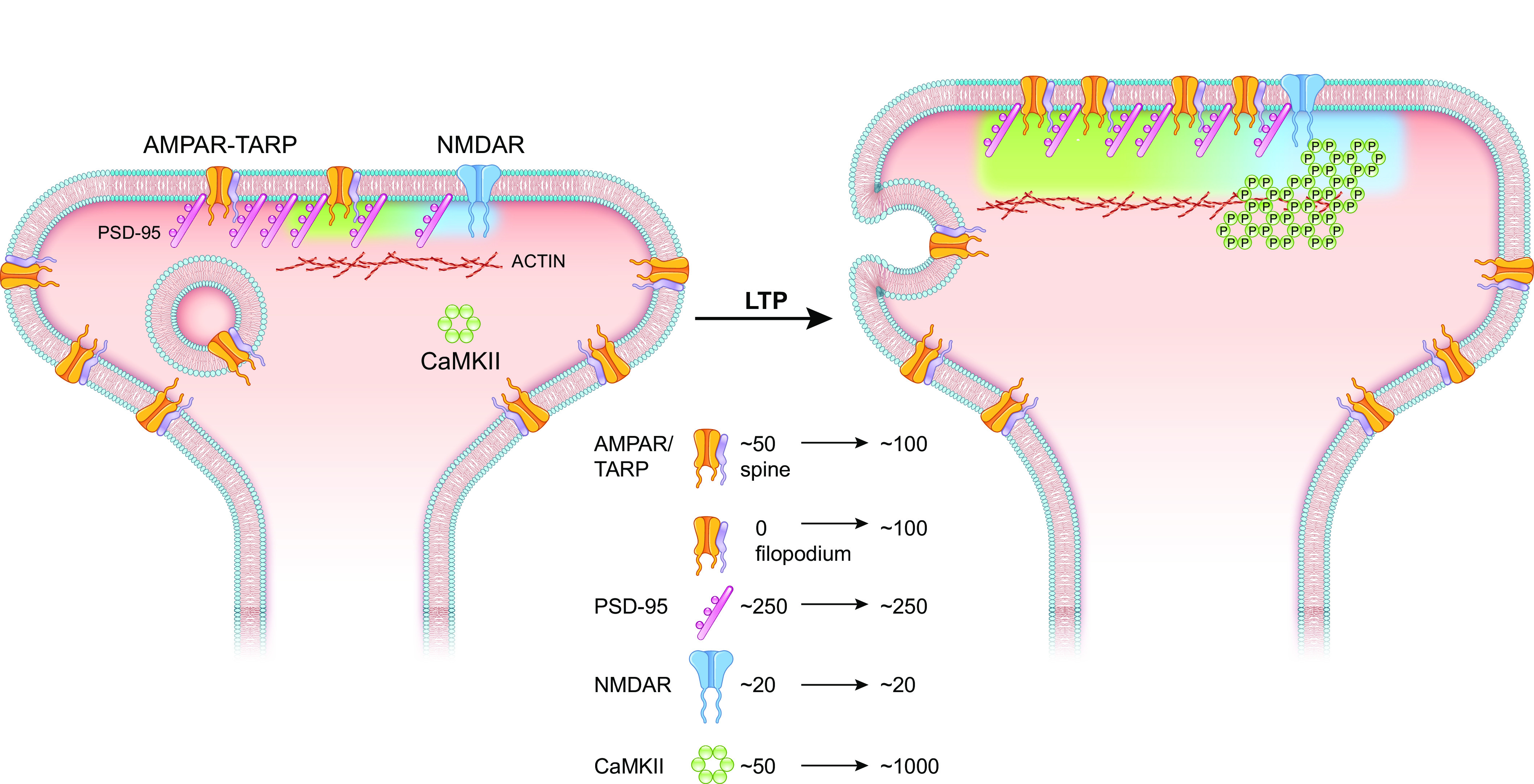
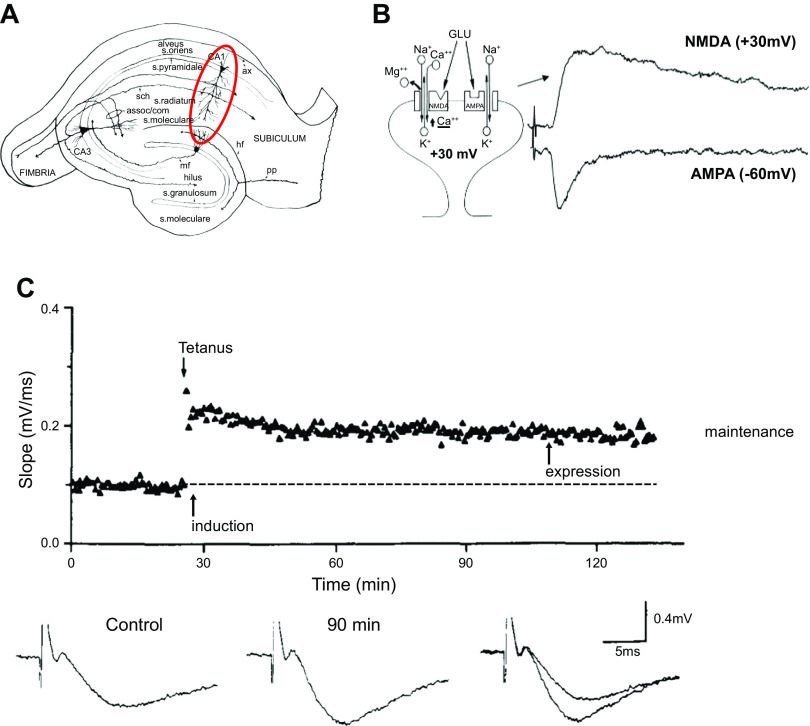
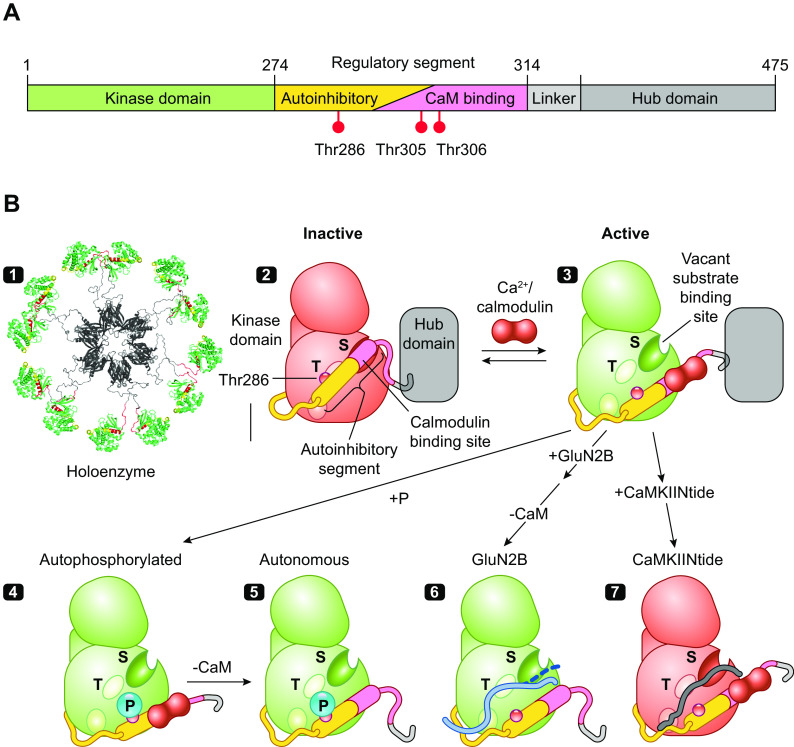
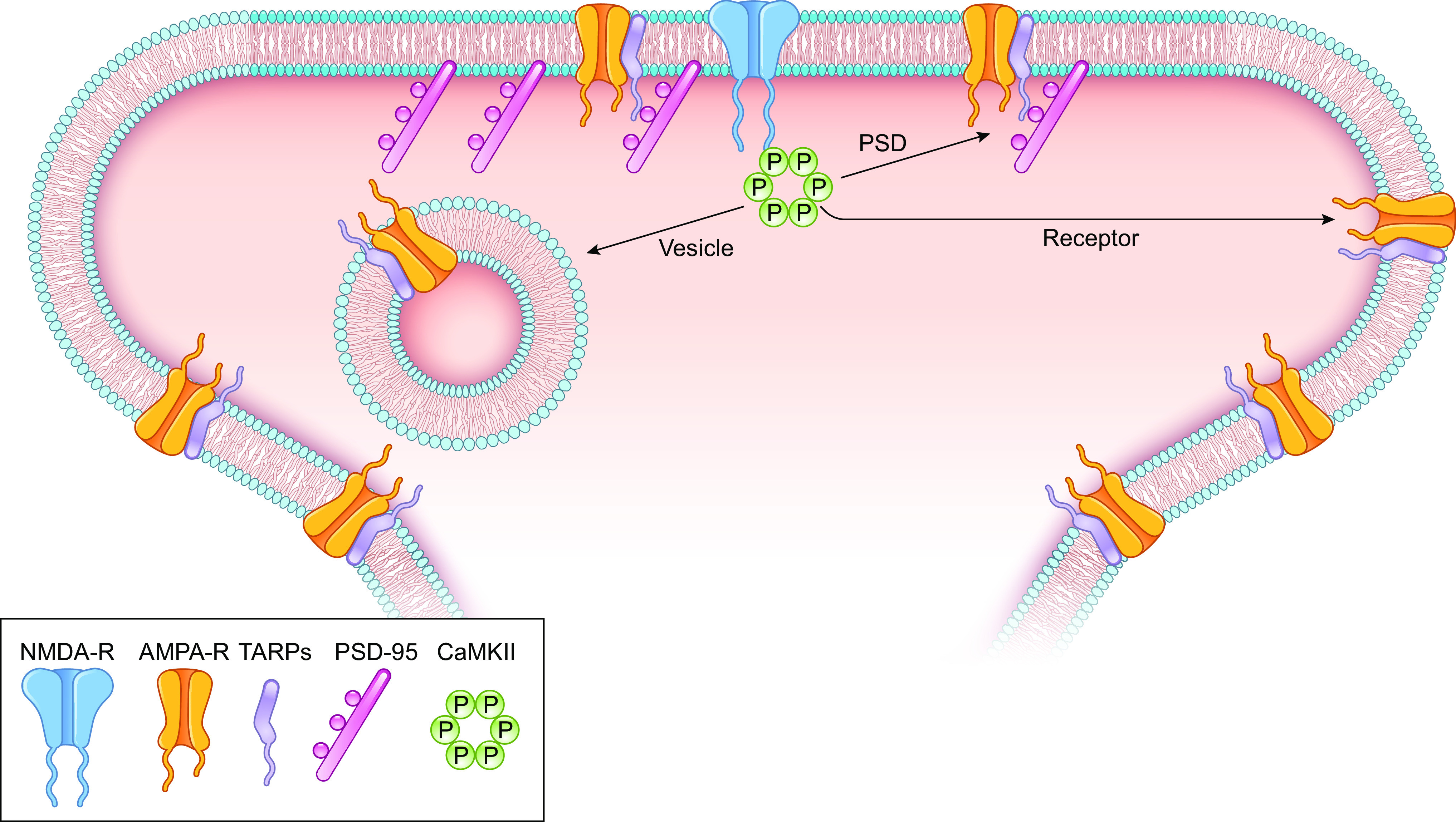
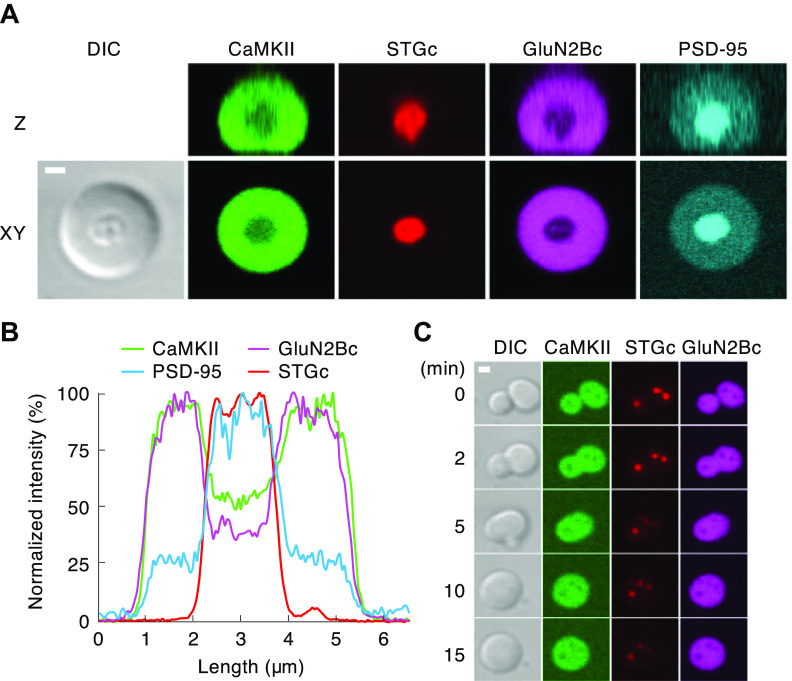
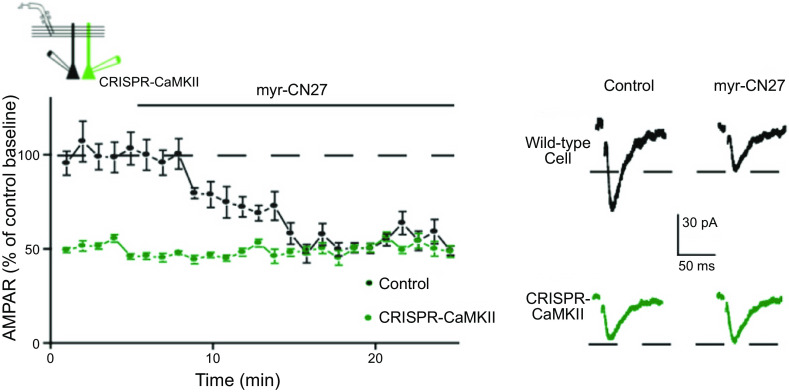
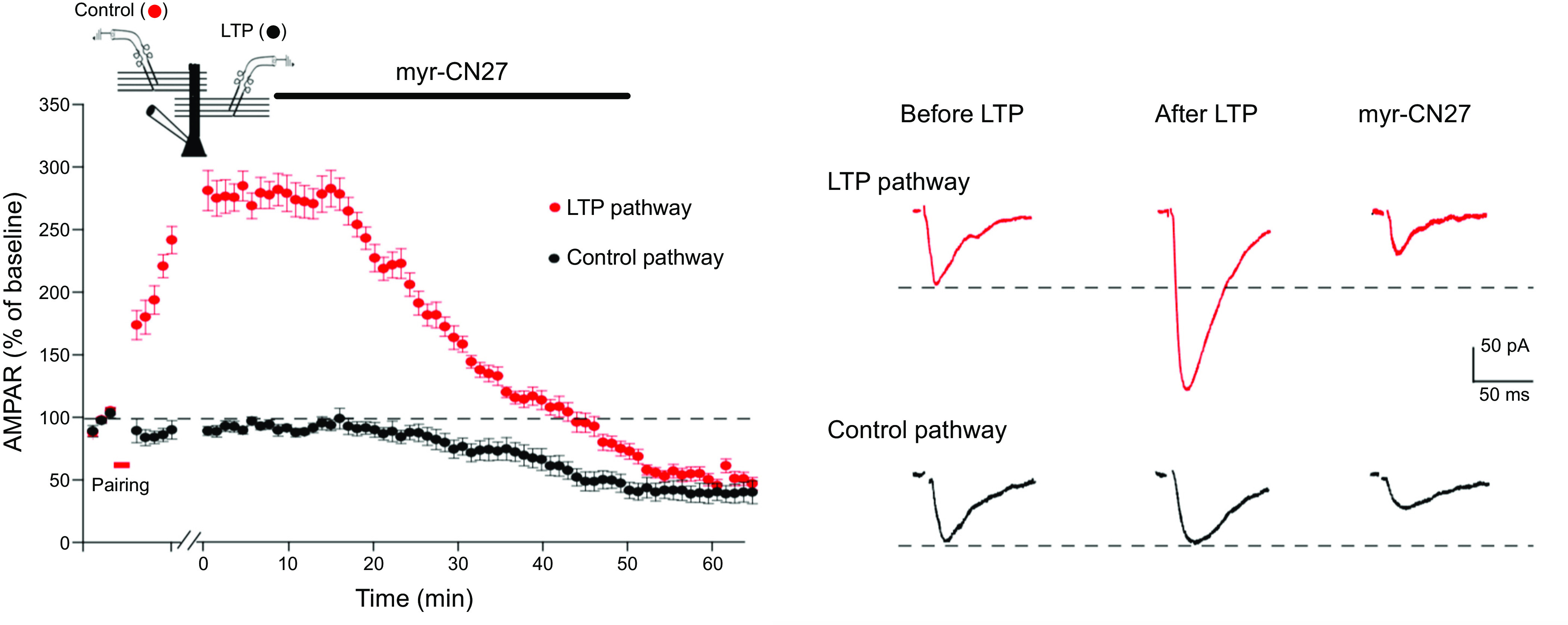
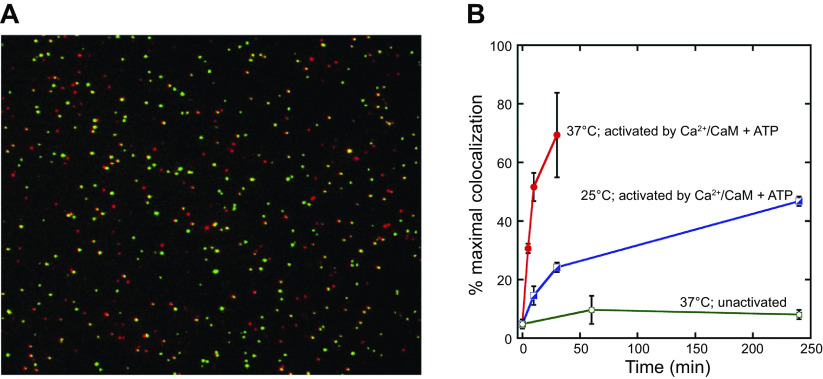
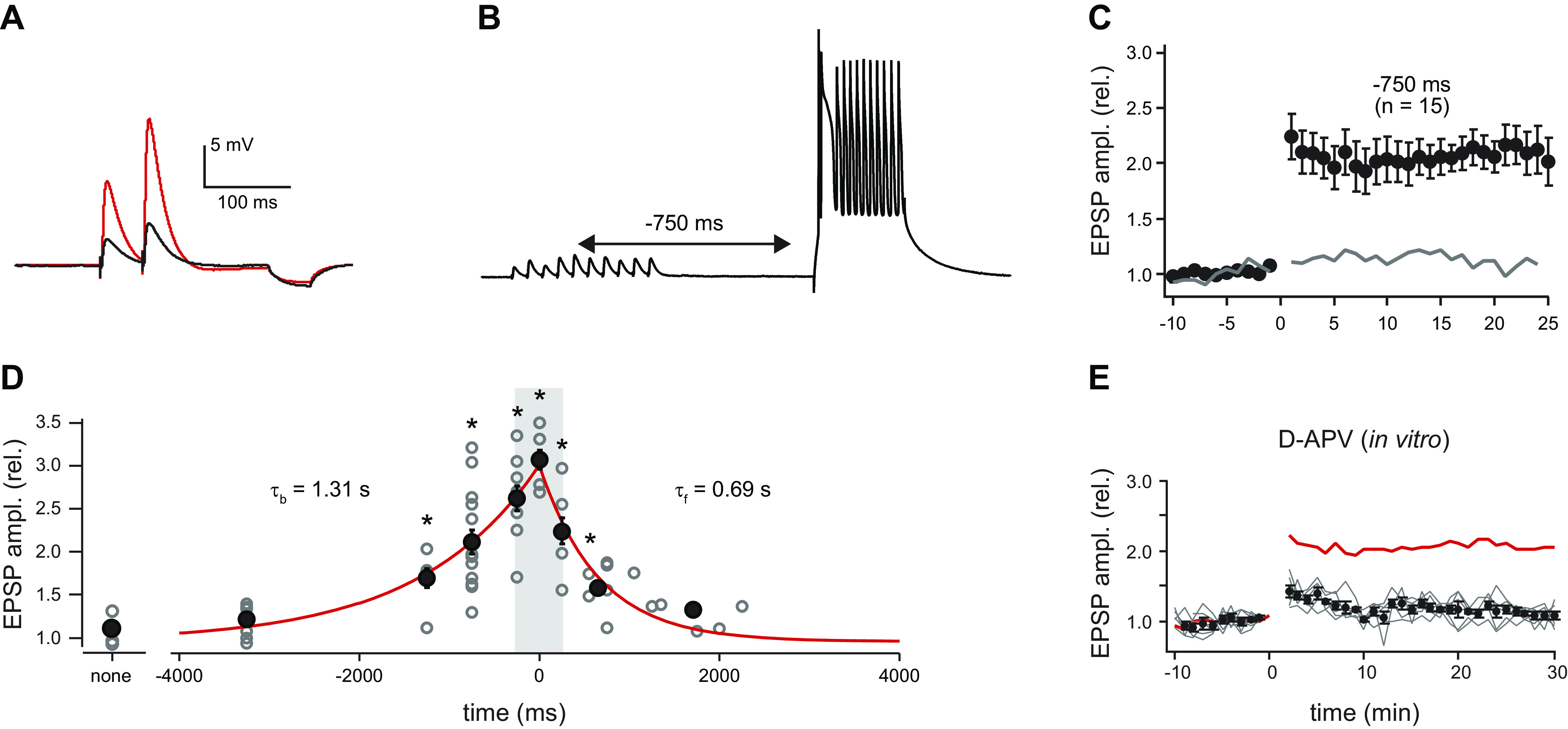
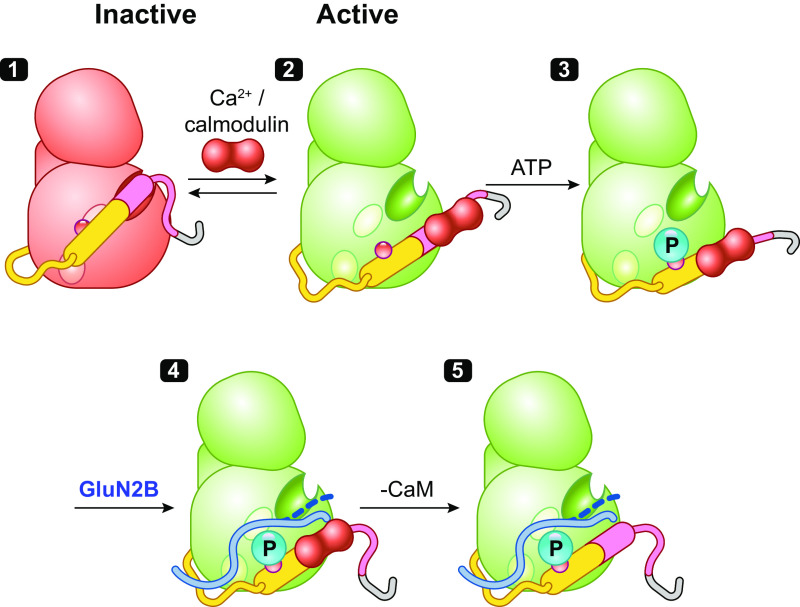
Ca/calmodulin-dependent protein kinase II (CaMKII) and long-term potentiation (LTP) were discovered within a decade of each other and have been inextricably intertwined ever since. However, like many marriages, it has had its up and downs. Based on the unique biochemical properties of CaMKII, it was proposed as a memory molecule before any physiological linkage was made to LTP. However, as reviewed here, the convincing linkage of CaMKII to synaptic physiology and behavior took many decades. New technologies were critical in this journey, including in vitro brain slices, mouse genetics, single-cell molecular genetics, pharmacological reagents, protein structure, and two-photon microscopy, as were new investigators attracted by the exciting challenge. This review tracks this journey and assesses the state of this marriage 40 years on. The collective literature impels us to propose a relatively simple model for synaptic memory involving the following steps that drive the process: ) Ca entry through -methyl-d-aspartate (NMDA) receptors activates CaMKII. ) CaMKII undergoes autophosphorylation resulting in constitutive, Ca-independent activity and exposure of a binding site for the NMDA receptor subunit GluN2B. ) Active CaMKII translocates to the postsynaptic density (PSD) and binds to the cytoplasmic C-tail of GluN2B. ) The CaMKII-GluN2B complex initiates a structural rearrangement of the PSD that may involve liquid-liquid phase separation. ) This rearrangement involves the PSD-95 scaffolding protein, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs), and their transmembrane AMPAR-regulatory protein (TARP) auxiliary subunits, resulting in an accumulation of AMPARs in the PSD that underlies synaptic potentiation. ) The stability of the modified PSD is maintained by the stability of the CaMKII-GluN2B complex. ) By a process of subunit exchange or interholoenzyme phosphorylation CaMKII maintains synaptic potentiation in the face of CaMKII protein turnover. There are many other important proteins that participate in enlargement of the synaptic spine or modulation of the steps that drive and maintain the potentiation. In this review we critically discuss the data underlying each of the steps. As will become clear, some of these steps are more firmly grounded than others, and we provide suggestions as to how the evidence supporting these steps can be strengthened or, based on the new data, be replaced. Although the journey has been a long one, the prospect of having a detailed cellular and molecular understanding of learning and memory is at hand.
钙/钙调蛋白依赖性蛋白激酶 II(CaMKII)和长时程增强(LTP)在彼此发现后的十年内被发现,并从此紧密交织在一起。然而,就像许多婚姻一样,它们也有起有落。基于 CaMKII 的独特生化特性,它在与 LTP 建立任何生理联系之前就被提出作为一种记忆分子。然而,正如本文所回顾的那样,将 CaMKII 与突触生理学和行为联系起来需要几十年的时间。新技术在这个过程中至关重要,包括体外脑片、小鼠遗传学、单细胞分子遗传学、药理学试剂、蛋白质结构和双光子显微镜,以及被这个令人兴奋的挑战吸引而来的新研究人员。本综述追踪了这段旅程,并评估了这个婚姻在 40 年后的现状。总的文献资料促使我们提出一个相对简单的突触记忆模型,涉及以下步骤,这些步骤推动了这个过程:1. 通过 N-甲基-D-天冬氨酸(NMDA)受体进入 Ca 会激活 CaMKII。2. CaMKII 发生自磷酸化,导致组成型、Ca 非依赖性活性和 NMDA 受体亚基 GluN2B 的结合位点暴露。3. 活性 CaMKII 易位到突触后密度(PSD)并与 GluN2B 的细胞质 C 尾巴结合。4. CaMKII-GluN2B 复合物引发 PSD 的结构重排,可能涉及液-液相分离。5. 这种重排涉及 PSD-95 支架蛋白、α-氨基-3-羟基-5-甲基-4-异恶唑丙酸受体(AMPAR)及其跨膜 AMPAR 调节蛋白(TARP)辅助亚基,导致 PSD 中 AMPAR 的积累,这是突触增强的基础。6. 通过亚基交换或同工酶磷酸化的过程,CaMKII 在 CaMKII 蛋白周转的情况下维持突触增强。还有许多其他重要的蛋白质参与到突触棘的扩大或驱动和维持增强的步骤的调节中。在这篇综述中,我们批判性地讨论了每个步骤的基础数据。显而易见的是,其中一些步骤比其他步骤更有根据,我们就如何加强支持这些步骤的证据或根据新数据替换它们提出了建议。尽管这段旅程漫长,但我们即将详细了解学习和记忆的细胞和分子基础。