Jiangsu Key Laboratory of Crop Genomics and Molecular Breeding/Key Laboratory of Plant Functional Genomics of the Ministry of Education, Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou, China.
State Key Lab of Plant Genomics, Institute of Genetics and Developmental Biology, Innovation Academy for Seed Design, Chinese Academy of Sciences, Beijing, China.
Elife. 2024 Jun 12;13:RP94180. doi: 10.7554/eLife.94180.
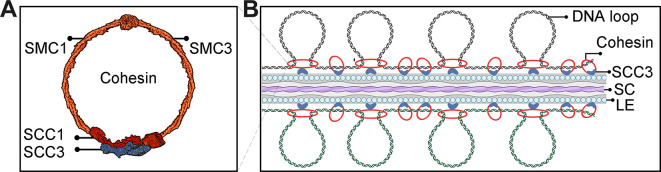
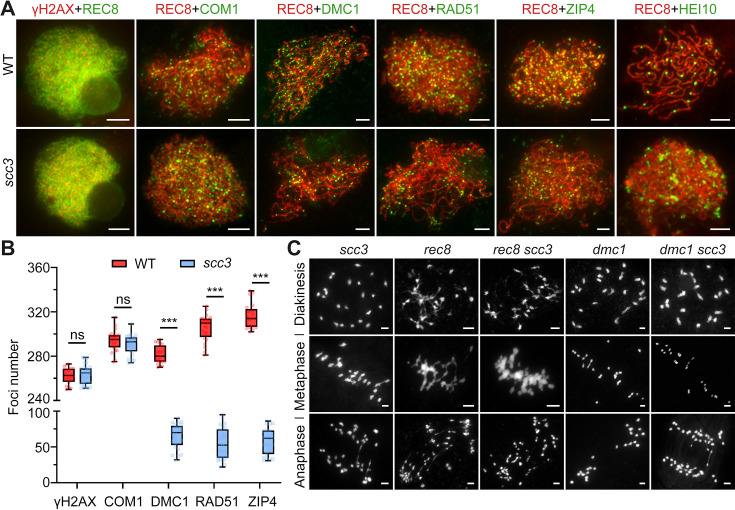
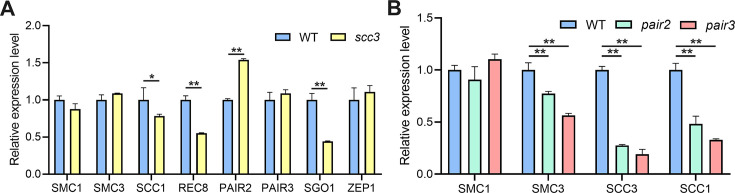
Cohesin is a multi-subunit protein that plays a pivotal role in holding sister chromatids together during cell division. Sister chromatid cohesion 3 (SCC3), constituents of cohesin complex, is highly conserved from yeast to mammals. Since the deletion of individual cohesin subunit always causes lethality, it is difficult to dissect its biological function in both mitosis and meiosis. Here, we obtained weak mutants using CRISPR-Cas9 system to explore its function during rice mitosis and meiosis. The weak mutants displayed obvious vegetative defects and complete sterility, underscoring the essential roles of SCC3 in both mitosis and meiosis. SCC3 is localized on chromatin from interphase to prometaphase in mitosis. However, in meiosis, SCC3 acts as an axial element during early prophase I and subsequently situates onto centromeric regions following the disassembly of the synaptonemal complex. The loading of SCC3 onto meiotic chromosomes depends on REC8. shows severe defects in homologous pairing and synapsis. Consequently, SCC3 functions as an axial element that is essential for maintaining homologous chromosome pairing and synapsis during meiosis.
着丝粒蛋白复合物由多个亚基组成,在有丝分裂和减数分裂过程中对于姐妹染色单体的黏合发挥着关键作用。从酵母到人,姐妹染色单体黏合蛋白 3(SCC3)作为着丝粒蛋白复合物的一个组成部分高度保守。由于单个着丝粒蛋白亚基的缺失往往会导致细胞死亡,因此很难在有丝分裂和减数分裂过程中对其生物学功能进行剖析。在这里,我们利用 CRISPR-Cas9 系统获得了 SCC3 的弱突变体,以探索其在水稻有丝分裂和减数分裂过程中的功能。SCC3 的弱突变体表现出明显的营养缺陷和完全不育的表型,这突出表明 SCC3 在有丝分裂和减数分裂中都具有重要作用。在有丝分裂中,SCC3 从间期到前期一直定位于染色质上。然而,在减数分裂中,SCC3 在早期的细线期充当着丝粒轴元件,随后在联会复合体解体后定位于着丝粒区域。SCC3 在减数分裂染色体上的加载依赖于 REC8。SCC3 的缺失导致同源配对和联会严重缺陷。因此,SCC3 作为着丝粒轴元件,对于维持减数分裂过程中同源染色体的配对和联会是必需的。