Advanced Water Management Centre, The University of Queensland, Brisbane, Queensland, Australia.
BMC Microbiol. 2010 Apr 1;10:98. doi: 10.1186/1471-2180-10-98.
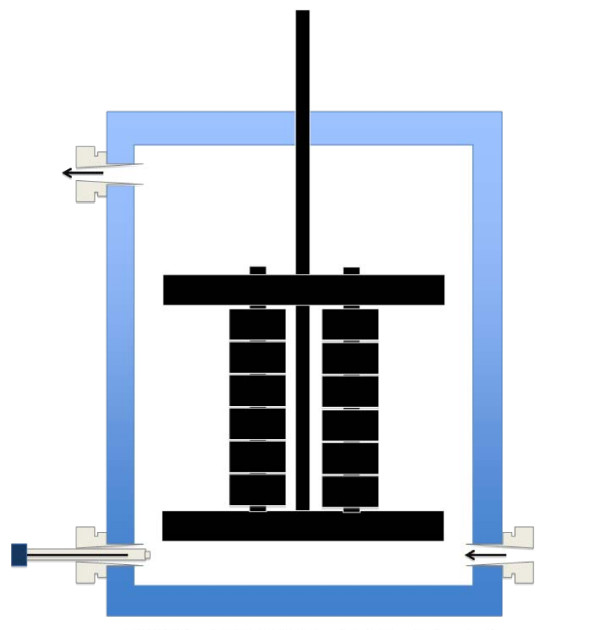
Microbial fuel cells (MFCs) rely on electrochemically active bacteria to capture the chemical energy contained in organics and convert it to electrical energy. Bacteria develop biofilms on the MFC electrodes, allowing considerable conversion capacity and opportunities for extracellular electron transfer (EET). The present knowledge on EET is centred around two Gram-negative models, i.e. Shewanella and Geobacter species, as it is believed that Gram-positives cannot perform EET by themselves as the Gram-negatives can. To understand how bacteria form biofilms within MFCs and how their development, structure and viability affects electron transfer, we performed pure and co-culture experiments.
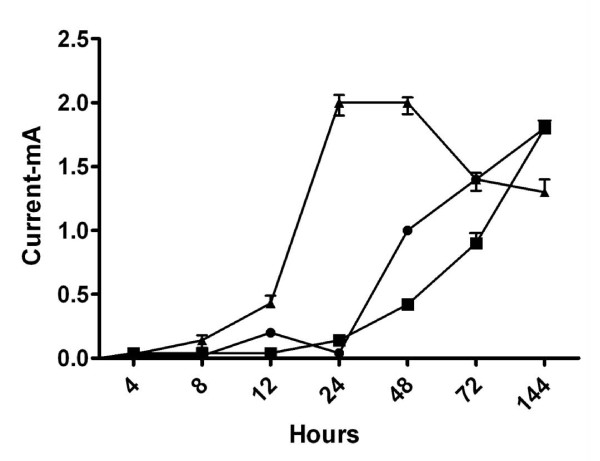
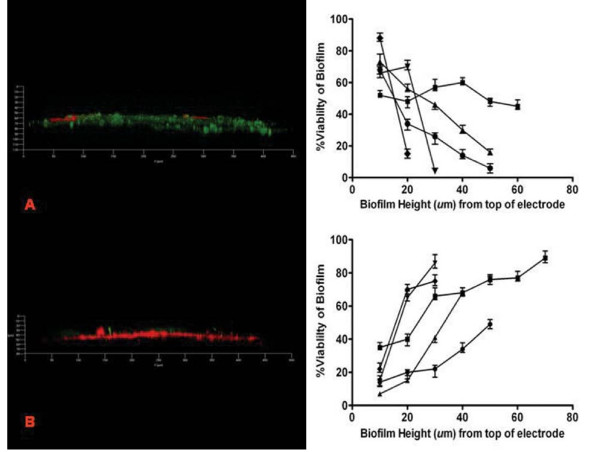
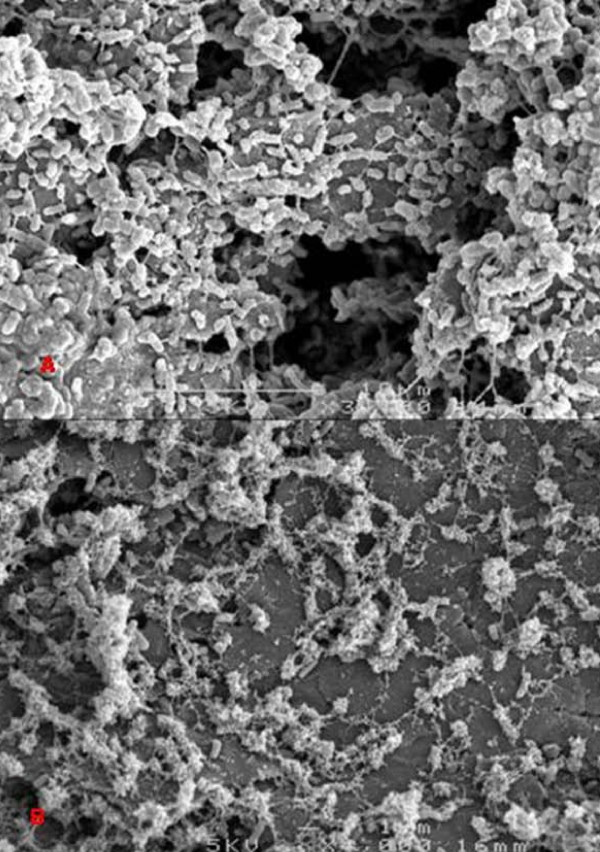
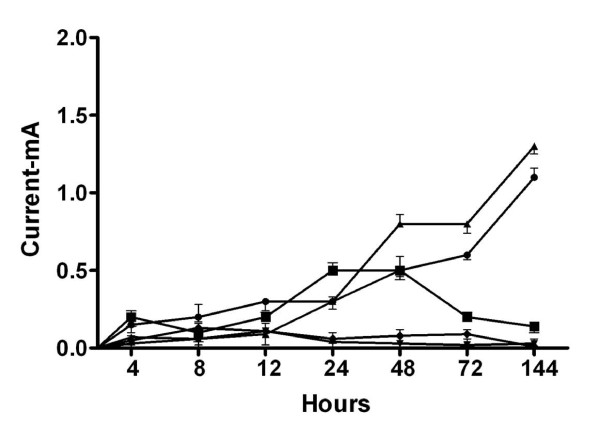
Biofilm viability was maintained highest nearer the anode during closed circuit operation (current flowing), in contrast to when the anode was in open circuit (soluble electron acceptor) where viability was highest on top of the biofilm, furthest from the anode. Closed circuit anode Pseudomonas aeruginosa biofilms were considerably thinner compared to the open circuit anode (30 +/- 3 microm and 42 +/- 3 microm respectively), which is likely due to the higher energetic gain of soluble electron acceptors used. The two Gram-positive bacteria used only provided a fraction of current produced by the Gram-negative organisms. Power output of co-cultures Gram-positive Enterococcus faecium and either Gram-negative organisms, increased by 30-70% relative to the single cultures. Over time the co-culture biofilms segregated, in particular, Pseudomonas aeruginosa creating towers piercing through a thin, uniform layer of Enterococcus faecium. P. aeruginosa and E. faecium together generated a current of 1.8 +/- 0.4 mA while alone they produced 0.9 +/- 0.01 and 0.2 +/- 0.05 mA respectively.
We postulate that this segregation may be an essential difference in strategy for electron transfer and substrate capture between the Gram-negative and the Gram-positive bacteria used here.
微生物燃料电池(MFC)依赖于电化学活性细菌来捕获有机物中所含的化学能,并将其转化为电能。细菌在 MFC 电极上形成生物膜,从而实现可观的转化能力和细胞外电子转移(EET)的机会。目前,关于 EET 的知识主要集中在两种革兰氏阴性模型,即希瓦氏菌和 Geobacter 属,因为人们认为革兰氏阳性菌不能像革兰氏阴性菌那样自行进行 EET。为了了解细菌如何在 MFC 中形成生物膜,以及它们的发育、结构和活力如何影响电子转移,我们进行了纯培养和共培养实验。
闭路操作(电流流动)时,靠近阳极的生物膜活力保持最高,而在阳极开路(可溶性电子受体)时,生物膜顶部(离阳极最远)的活力最高。与阳极开路相比,闭路阳极铜绿假单胞菌生物膜要薄得多(分别为 30 ± 3 微米和 42 ± 3 微米),这可能是由于使用了更高能量的可溶性电子受体。所使用的两种革兰氏阳性菌仅提供了革兰氏阴性菌产生电流的一部分。与单一培养物相比,共培养革兰氏阳性粪肠球菌和革兰氏阴性菌的组合,其功率输出增加了 30-70%。随着时间的推移,共培养生物膜发生了分离,特别是铜绿假单胞菌形成了刺穿粪肠球菌薄而均匀层的塔。铜绿假单胞菌和粪肠球菌一起产生 1.8 ± 0.4 mA 的电流,而单独使用时,它们分别产生 0.9 ± 0.01 和 0.2 ± 0.05 mA 的电流。
我们推测,这种分离可能是这里使用的革兰氏阴性和革兰氏阳性菌在电子转移和底物捕获方面的策略的一个重要区别。