Department of Biochemistry, 89 Beaumont Avenue, Burlington, VT 05405, USA.
Biochemistry. 2010 Dec 7;49(48):10329-38. doi: 10.1021/bi101130t. Epub 2010 Nov 10.
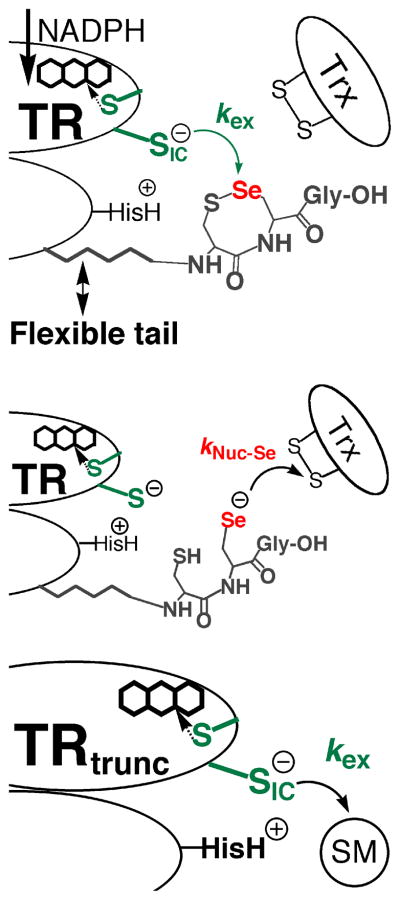
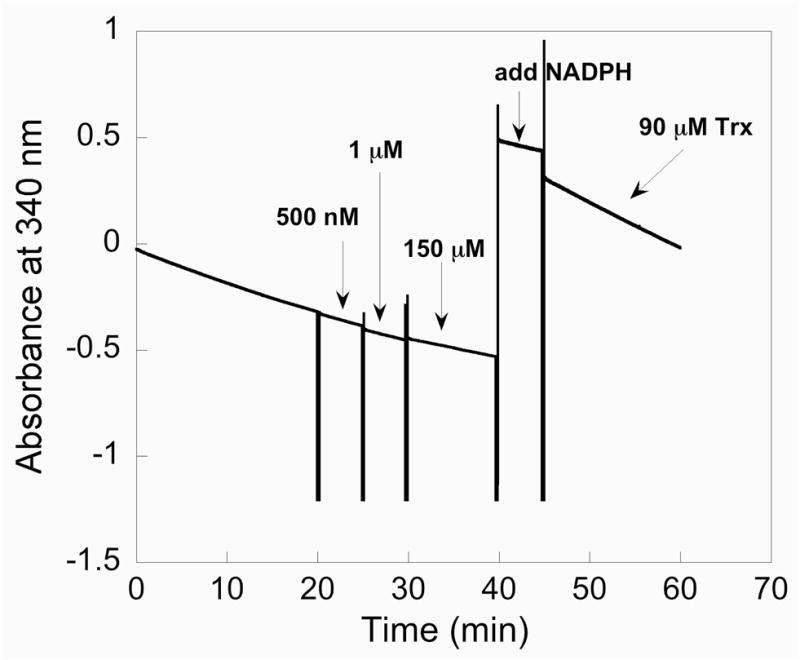
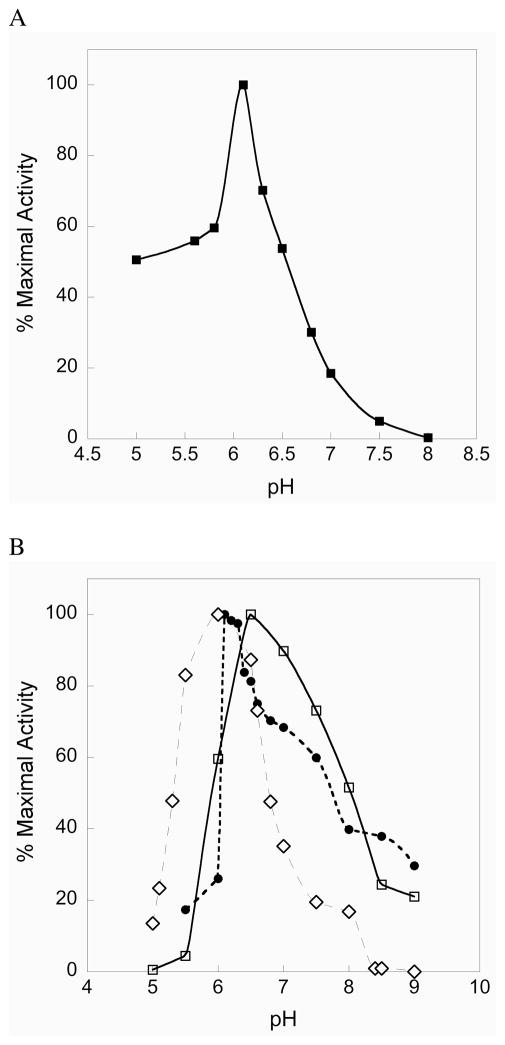
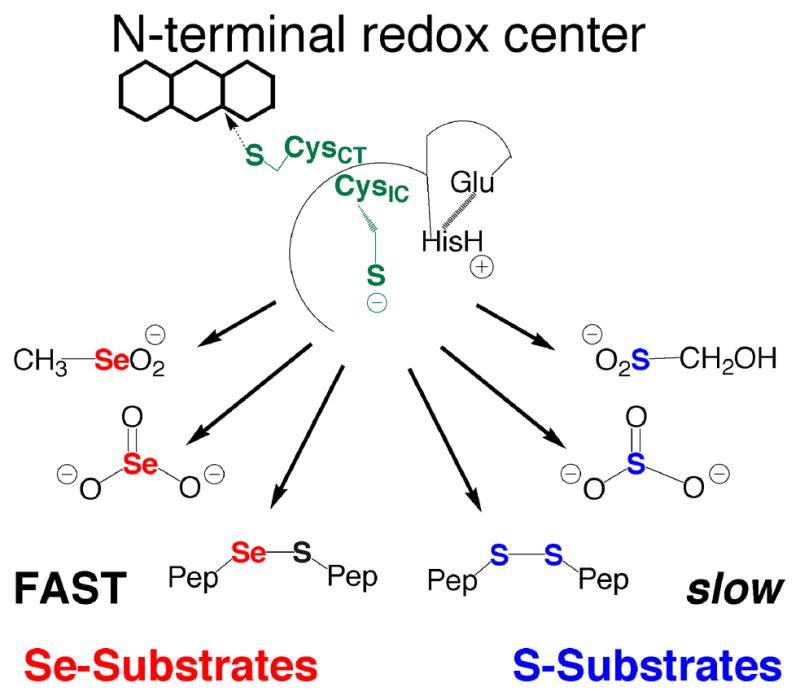
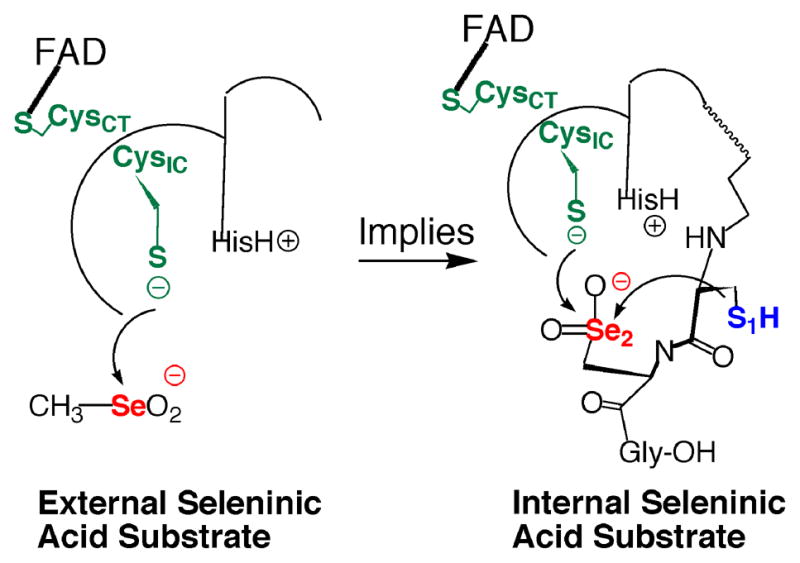
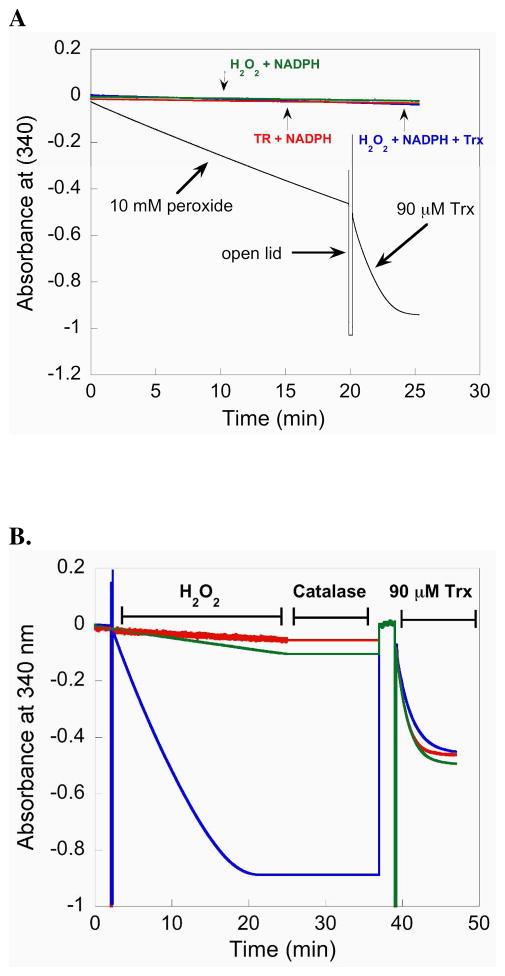
Mammalian thioredoxin reductase is a homodimeric pyridine nucleotide disulfide oxidoreductase that contains the rare amino acid selenocysteine (Sec) on a C-terminal extension. We previously have shown that a truncated version of mouse mitochondrial thioredoxin reductase missing this C-terminal tail will catalyze the reduction of a number of small molecules. Here we show that the truncated thioredoxin reductase will catalyze the reduction of methaneseleninic acid. This reduction is fast at pH 6.1 and is only 4-fold slower than that of the full-length enzyme containing Sec. This finding suggested to us that if the C-terminal Sec residue in the holoenzyme became oxidized to the seleninic acid form (Sec-SeO(2)(-)) that it would be quickly reduced back to an active state by enzymic thiols and further suggested to us that the enzyme would be very resistant to irreversible inactivation by oxidation. We tested this hypothesis by reducing the enzyme with NADPH and subjecting it to high concentrations of H(2)O(2) (up to 50 mM). The results show that the enzyme strongly resisted inactivation by 50 mM H(2)O(2). To determine the redox state of the C-terminal Sec residue, we attempted to inhibit the enzyme with dimedone. Dimedone alkylates protein sulfenic acid residues and presumably will alkylate selenenic acid (Sec-SeOH) residues as well. The enzyme was not inhibited by dimedone even when a 150-fold excess was added to the reaction mixture containing the enzyme and H(2)O(2). We also tested the ability of the truncated enzyme to resist inactivation by oxidation as well and found that it also was resistant to high concentrations of H(2)O(2). One assumption for the use of Sec in enzymes is that it is catalytically superior to the use of cysteine. We and others have previously suggested that there are reasons for the use of Sec in enzymes that are unrelated to the conversion of substrate to product. The data presented here support this assertion. The results also imply that the redox signaling function of the thioredoxin system can remain active under oxidative stress.
哺乳动物硫氧还蛋白还原酶是一种同二聚体吡啶核苷酸二硫化物氧化还原酶,在 C 端延伸部分含有罕见的氨基酸硒代半胱氨酸(Sec)。我们之前已经表明,缺失这个 C 端尾部的小鼠线粒体硫氧还蛋白的截断版本将催化许多小分子的还原。在这里,我们表明,截断的硫氧还蛋白还原酶将催化甲硒酸的还原。在 pH6.1 时,这种还原反应非常快,仅比含有 Sec 的全长酶慢 4 倍。这一发现使我们认为,如果全酶中的 C 端 Sec 残基氧化为硒酸形式(Sec-SeO(2)(-)),那么它将被酶促硫醇迅速还原回活性状态,并进一步使我们认为该酶将非常抵抗因氧化而不可逆失活。我们通过用 NADPH 还原酶并使其暴露于高浓度 H(2)O(2)(高达 50mM)来测试这一假设。结果表明,该酶强烈抵抗 50mM H(2)O(2)的失活。为了确定 C 端 Sec 残基的氧化还原状态,我们尝试用二甲基酮抑制酶。二甲基酮烷基化蛋白质亚磺酰基残基,并且大概也会烷基化硒酸(Sec-SeOH)残基。即使在含有酶和 H(2)O(2)的反应混合物中加入 150 倍过量的二甲基酮,酶也没有被抑制。我们还测试了截断酶抵抗氧化失活的能力,发现它也抵抗高浓度的 H(2)O(2)。在酶中使用 Sec 的一个假设是它在催化上优于使用半胱氨酸。我们和其他人之前曾提出,在酶中使用 Sec 有与将底物转化为产物无关的原因。这里呈现的数据支持这一说法。结果还暗示,硫氧还蛋白系统的氧化还原信号功能可以在氧化应激下保持活跃。