Geeven Geert, Zhu Yun, Kim Byung Ju, Bartholdy Boris A, Yang Seung-Min, Macfarlan Todd S, Gifford Wesley D, Pfaff Samuel L, Verstegen Marjon J A M, Pinto Hugo, Vermunt Marit W, Creyghton Menno P, Wijchers Patrick J, Stamatoyannopoulos John A, Skoultchi Arthur I, de Laat Wouter
Hubrecht Institute-KNAW & University Medical Center Utrecht, Uppsalalaan 8, 3584, CT, Utrecht, The Netherlands.
Department of Cell Biology, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY, 10461, USA.
Genome Biol. 2015 Dec 23;16:289. doi: 10.1186/s13059-015-0857-0.
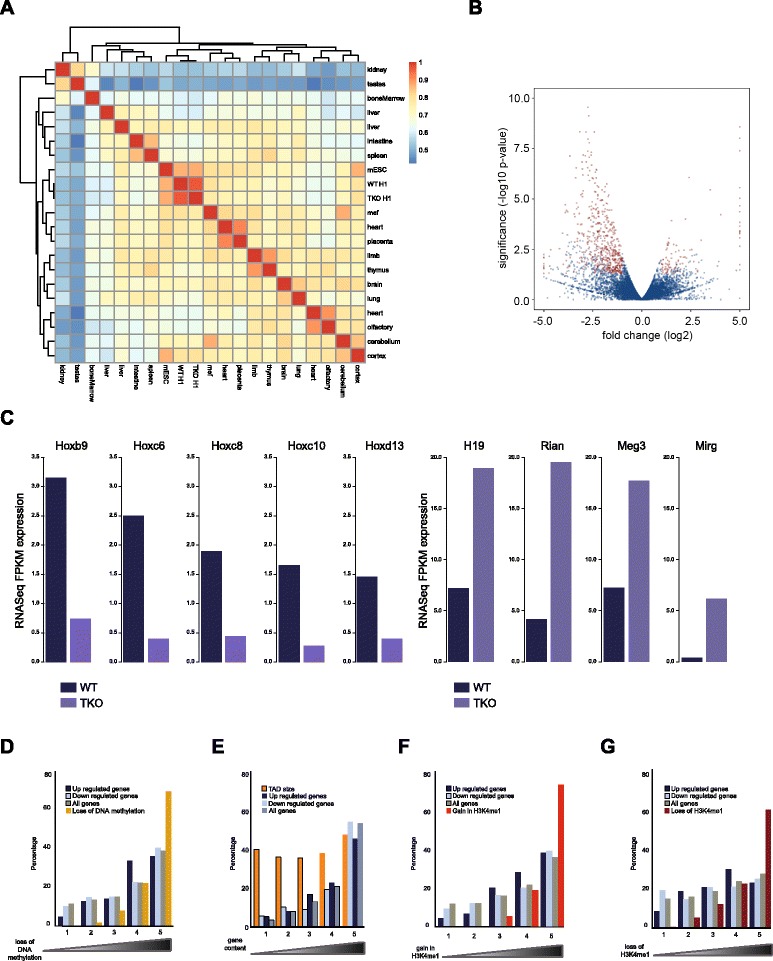
Linker histone H1 is a core chromatin component that binds to nucleosome core particles and the linker DNA between nucleosomes. It has been implicated in chromatin compaction and gene regulation and is anticipated to play a role in higher-order genome structure. Here we have used a combination of genome-wide approaches including DNA methylation, histone modification and DNase I hypersensitivity profiling as well as Hi-C to investigate the impact of reduced cellular levels of histone H1 in embryonic stem cells on chromatin folding and function.
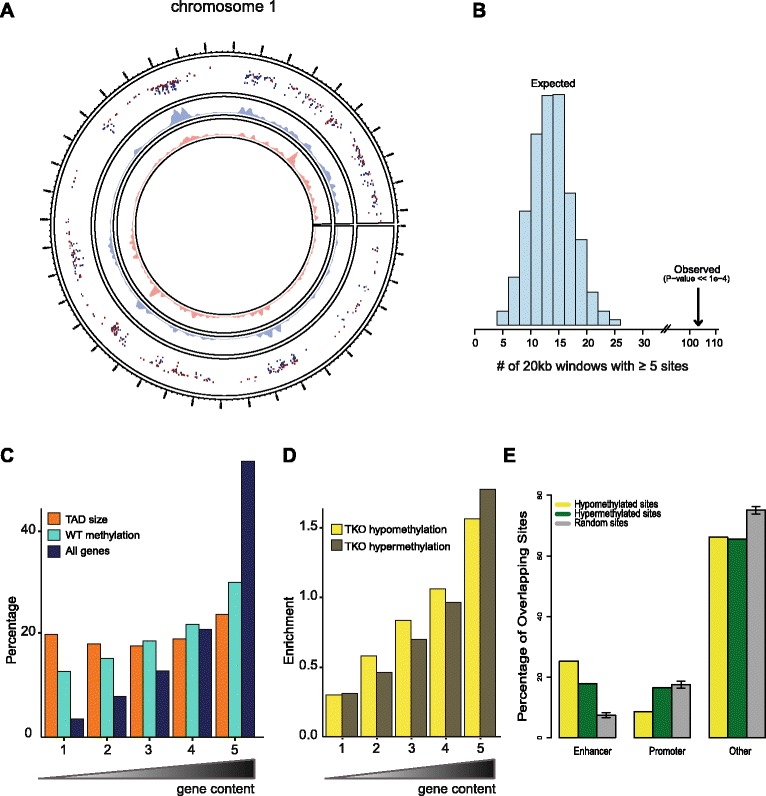
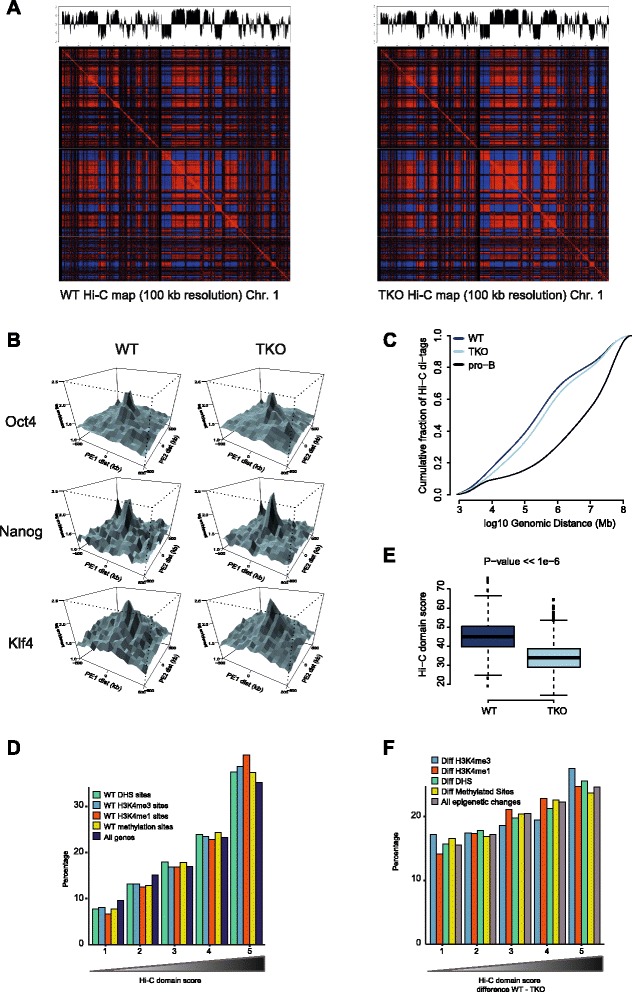
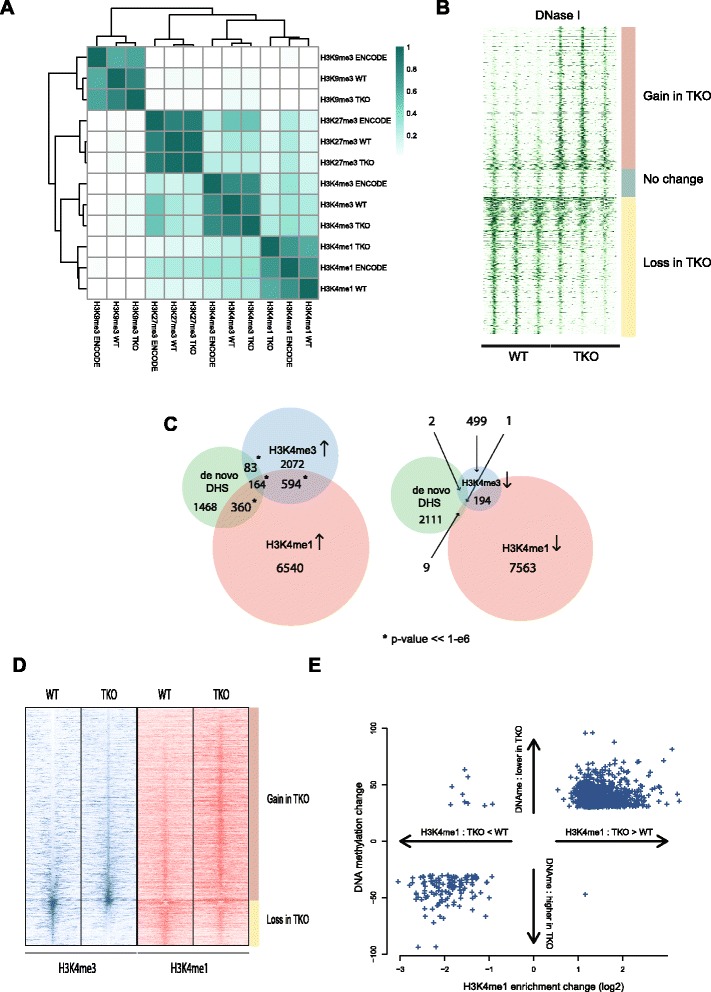
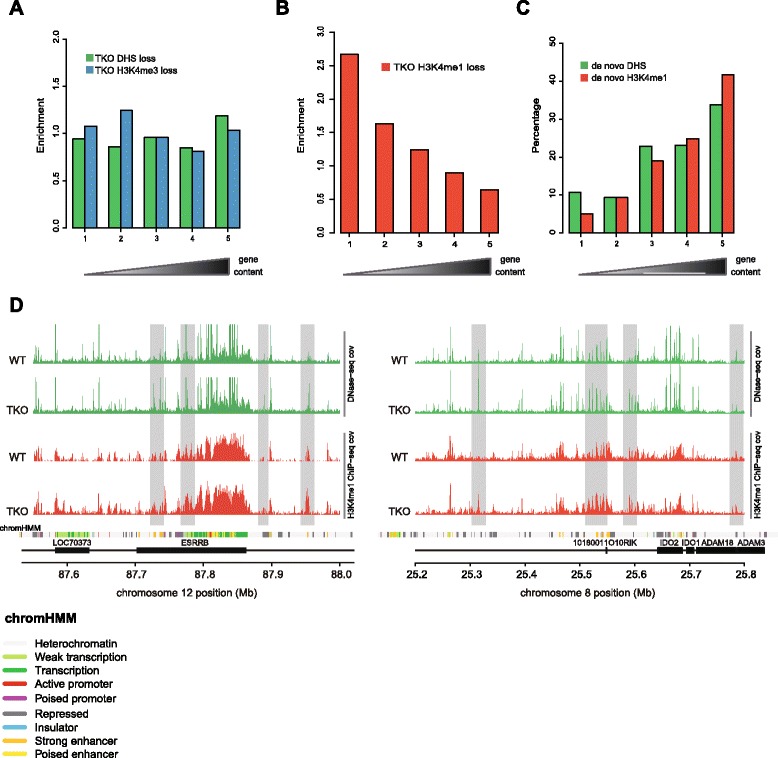
We find that depletion of histone H1 changes the epigenetic signature of thousands of potential regulatory sites across the genome. Many of them show cooperative loss or gain of multiple chromatin marks. Epigenetic alterations cluster to gene-dense topologically associating domains (TADs) that already showed a high density of corresponding chromatin features. Genome organization at the three-dimensional level is largely intact, but we find changes in the structural segmentation of chromosomes specifically for the epigenetically most modified TADs.
Our data show that cells require normal histone H1 levels to expose their proper regulatory landscape. Reducing the levels of histone H1 results in massive epigenetic changes and altered topological organization particularly at the most active chromosomal domains. Changes in TAD configuration coincide with epigenetic landscape changes but not with transcriptional output changes, supporting the emerging concept that transcriptional control and nuclear positioning of TADs are not causally related but independently controlled by the locally associated trans-acting factors.
连接组蛋白H1是一种核心染色质成分,它与核小体核心颗粒以及核小体之间的连接DNA相结合。它与染色质压缩和基因调控有关,并预计在高阶基因组结构中发挥作用。在这里,我们使用了包括DNA甲基化、组蛋白修饰和DNase I超敏反应谱分析以及Hi-C在内的全基因组方法组合,来研究胚胎干细胞中组蛋白H1细胞水平降低对染色质折叠和功能的影响。
我们发现组蛋白H1的缺失改变了全基因组数千个潜在调控位点的表观遗传特征。其中许多位点显示出多种染色质标记的协同缺失或增加。表观遗传改变聚集在已经显示出高密度相应染色质特征的基因密集型拓扑相关结构域(TAD)中。三维水平的基因组组织在很大程度上是完整的,但我们发现染色体的结构分割发生了变化,特别是对于表观遗传修饰最多的TAD。
我们的数据表明,细胞需要正常的组蛋白H1水平来展现其适当的调控格局。降低组蛋白H1的水平会导致大量表观遗传变化和拓扑组织改变,特别是在最活跃的染色体结构域。TAD构型的变化与表观遗传格局的变化一致,但与转录输出的变化不一致,这支持了一个新出现的概念,即TAD的转录控制和核定位不是因果相关的,而是由局部相关的反式作用因子独立控制的。